
神经细胞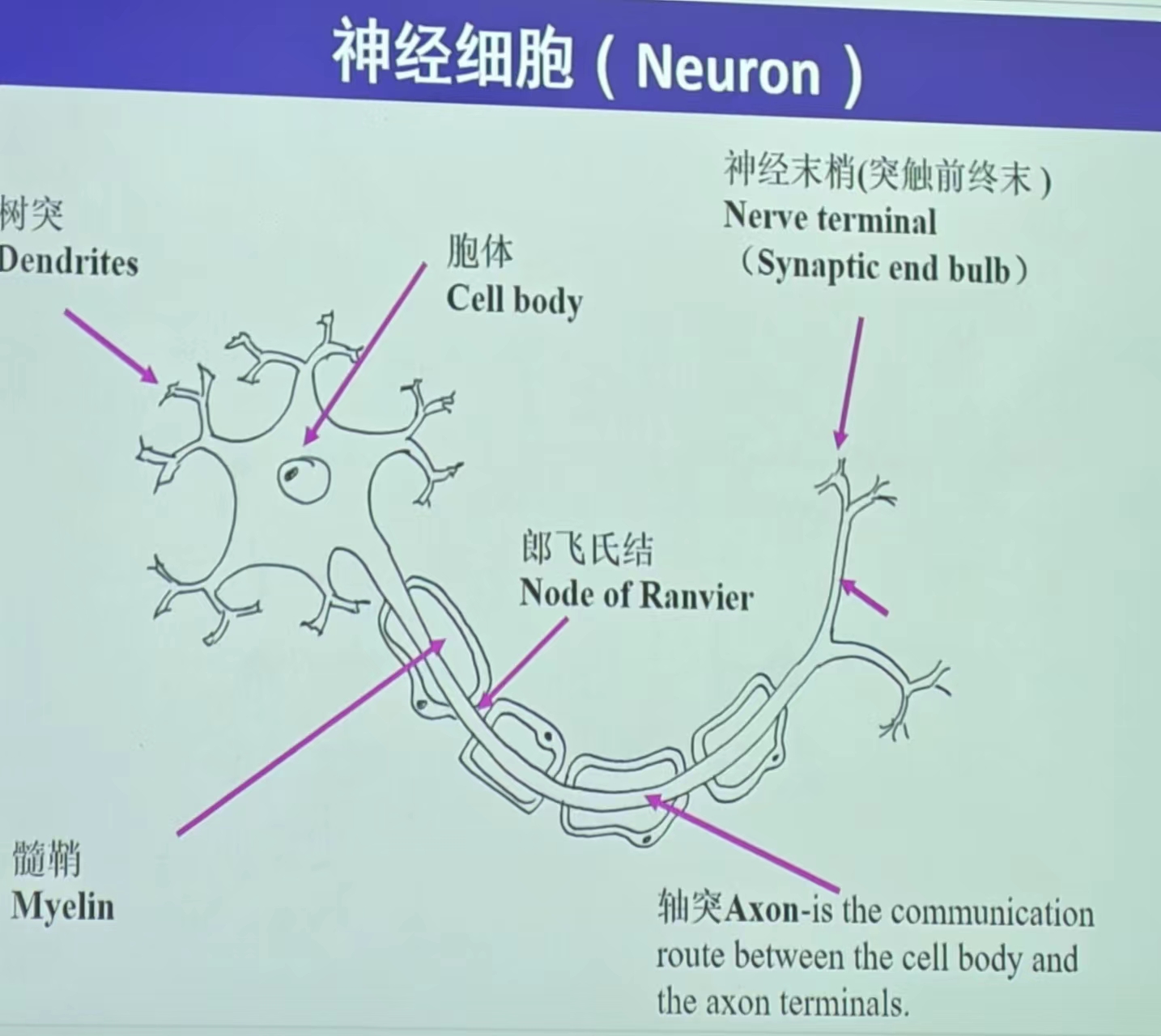
- 神经的电特性 ^ba12e0
- 历史脉络:(1)Galvani通过蛙的坐骨神经发现生物电
 (2)通过万用表可验证神经细胞的电势差
(2)通过万用表可验证神经细胞的电势差 - 静息电位的机制:细胞膜类似半透膜(Berstein),使得离子浓度处于动态平衡的状态(离子通道的存在?A.H—膜片钳的发现验证了A.H.的理论)

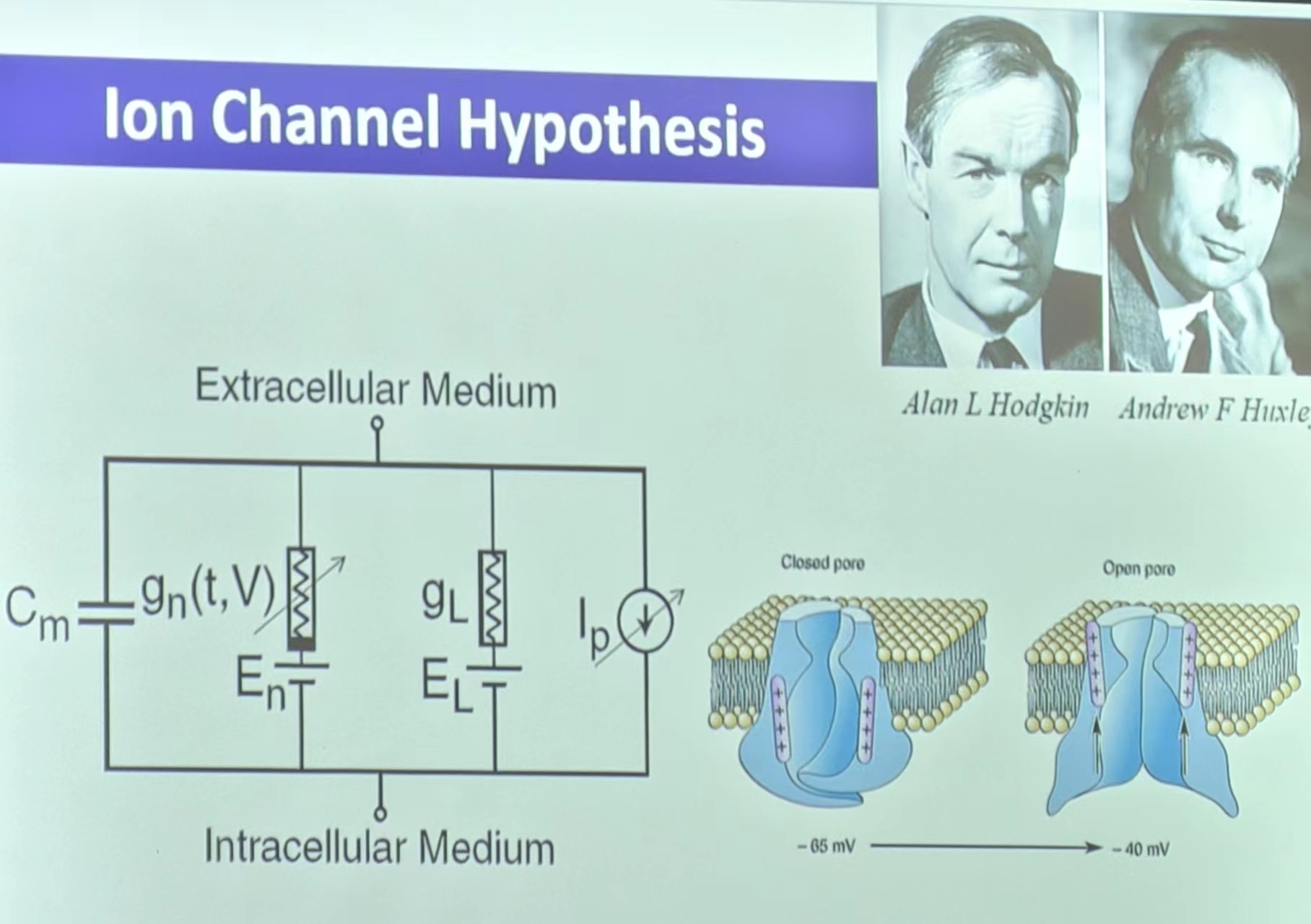
 注:电环路机制如下(解释看下文)
注:电环路机制如下(解释看下文)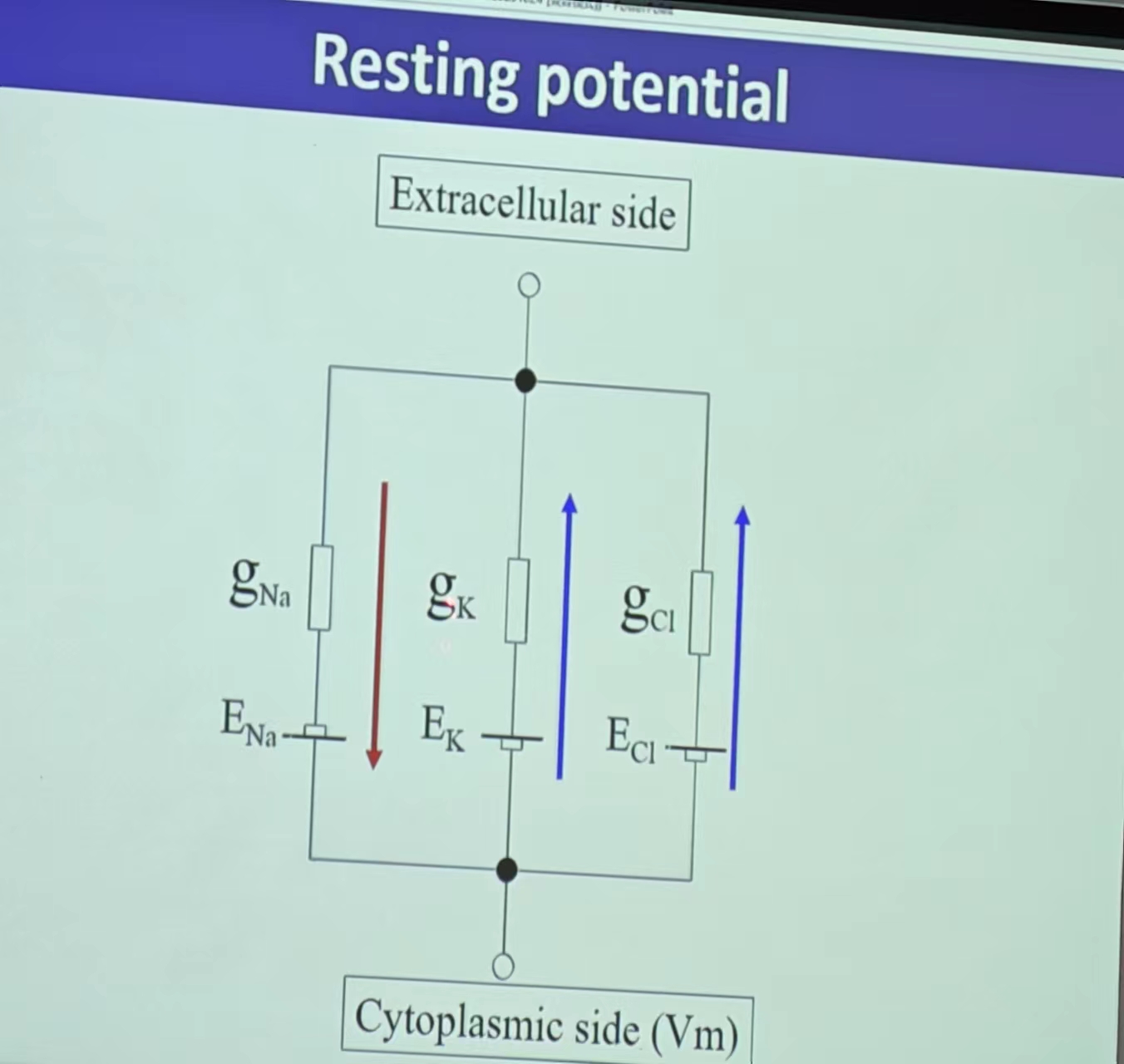
- 离子通道(电学角度:楞次电压【电压参考细胞外相对于细胞内的电压】串联电阻):

-
- 四大特点:pores、passive、selective (如Na离子结合水之后无法通过通道、离子通道里专门有识别的位点)、open【gating】 (能控—受体配体结合门控、磷酸化门控、电压门控、机械张力门控) /close—超过阈值只是打开或者关闭的可能性提高 (all or none——非绝对打开);不需ATP
-
- Rectifying(整形) Channel: gating时,其不完全符合欧姆定律,如NMDAR受体受Mg离子阻断的影响

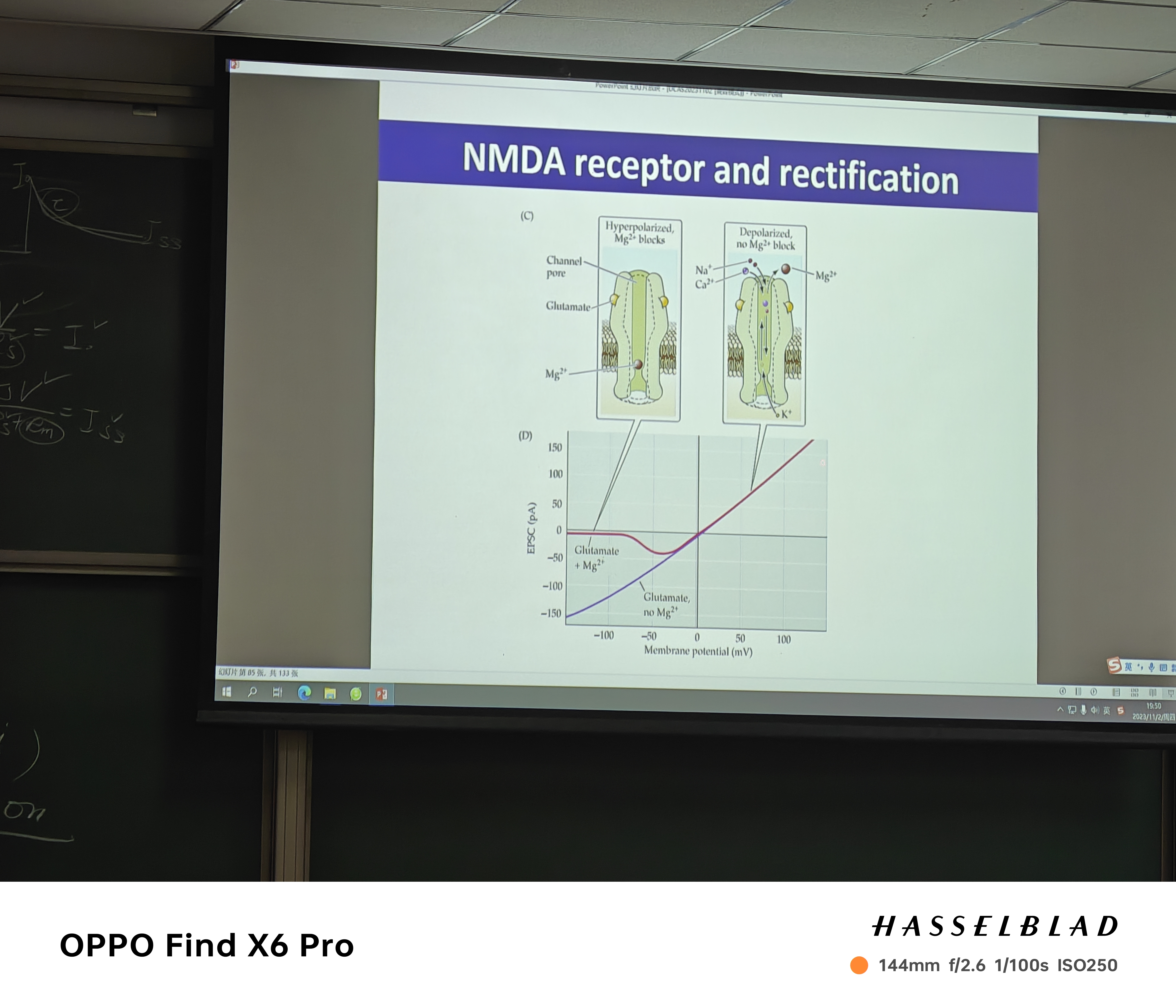
- Rectifying(整形) Channel: gating时,其不完全符合欧姆定律,如NMDAR受体受Mg离子阻断的影响
-
- All or none—probability
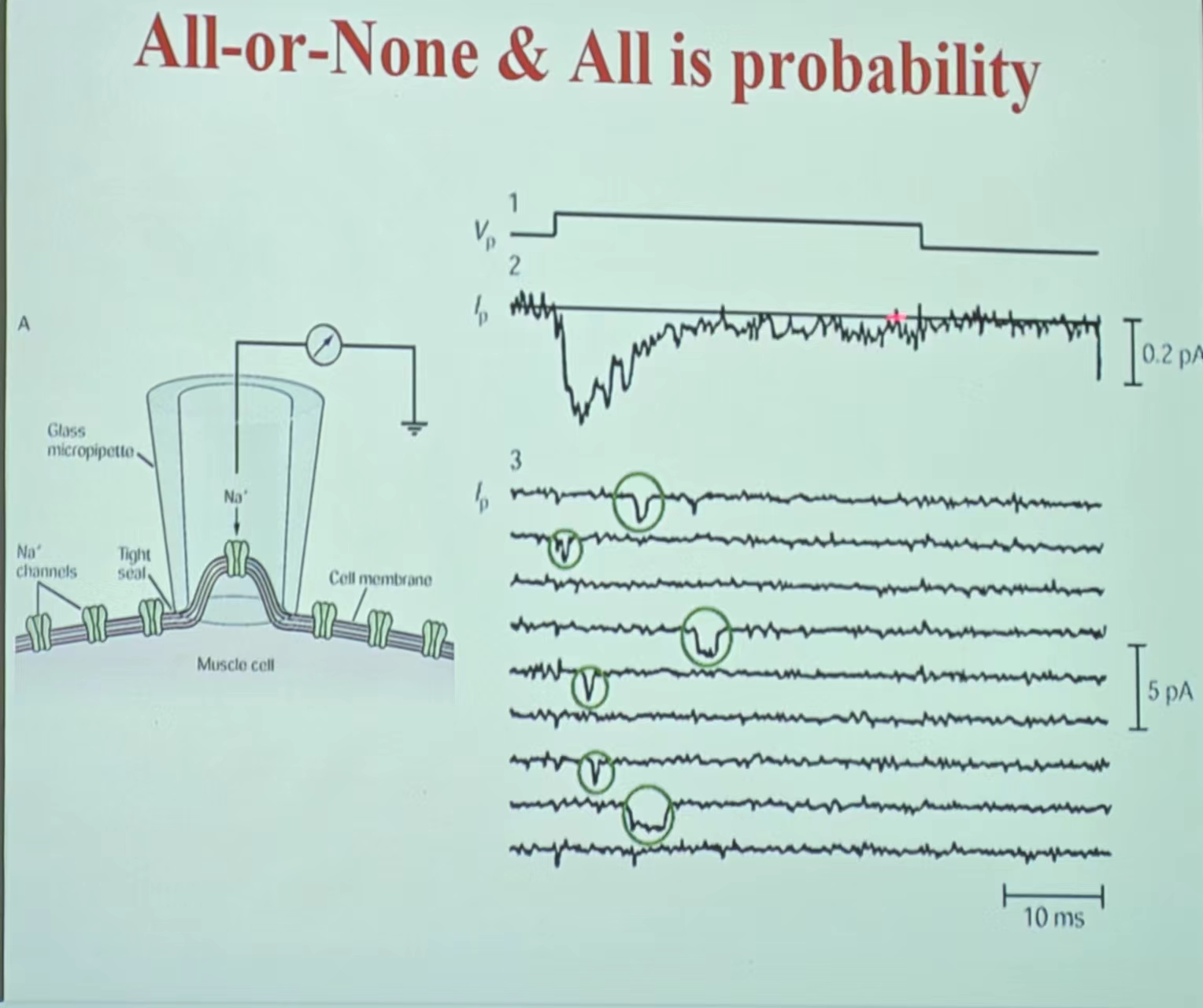
- All or none—probability
- 离子运动:受电场力(Potential)和diffusion force(高浓度向低浓度的运动)的影响——产生micro/macro force的平衡(动态平衡 Ix=0 【but:电位高于平衡电位,为正,正为外向电流,即胞内向胞外,反之。But电流不等于离子流,外向电流是阳离子的离子流,内向电流是阴离子的离子流 (阴离子内流,其介导的电流是往外流,反之) 】—楞次定律)—E:某种离子的平衡电位(Z带±)

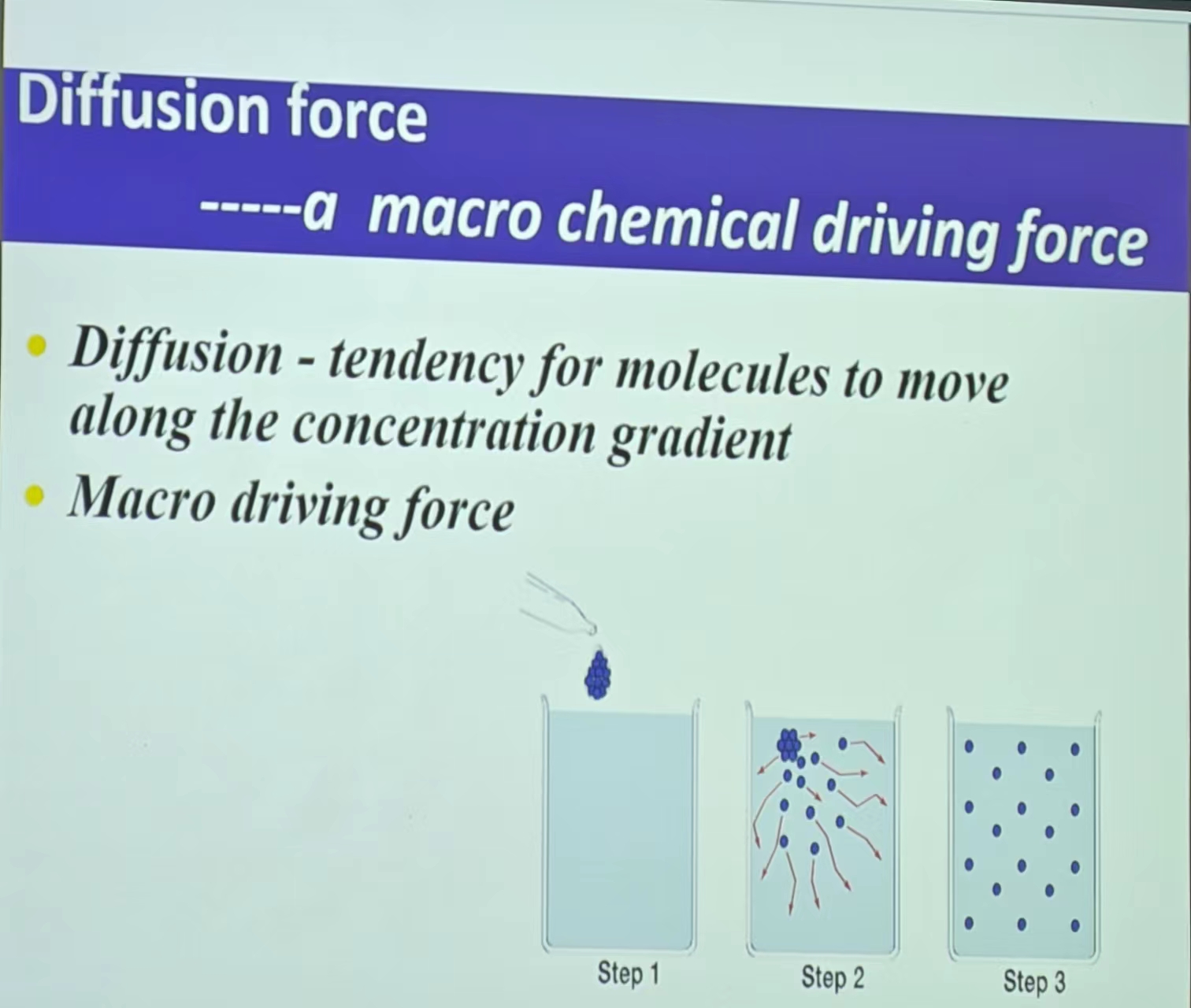 注:电场力和扩散力的离子流平衡如下,Ex见静息电位机制:
注:电场力和扩散力的离子流平衡如下,Ex见静息电位机制:

- 离子平衡电位例题:
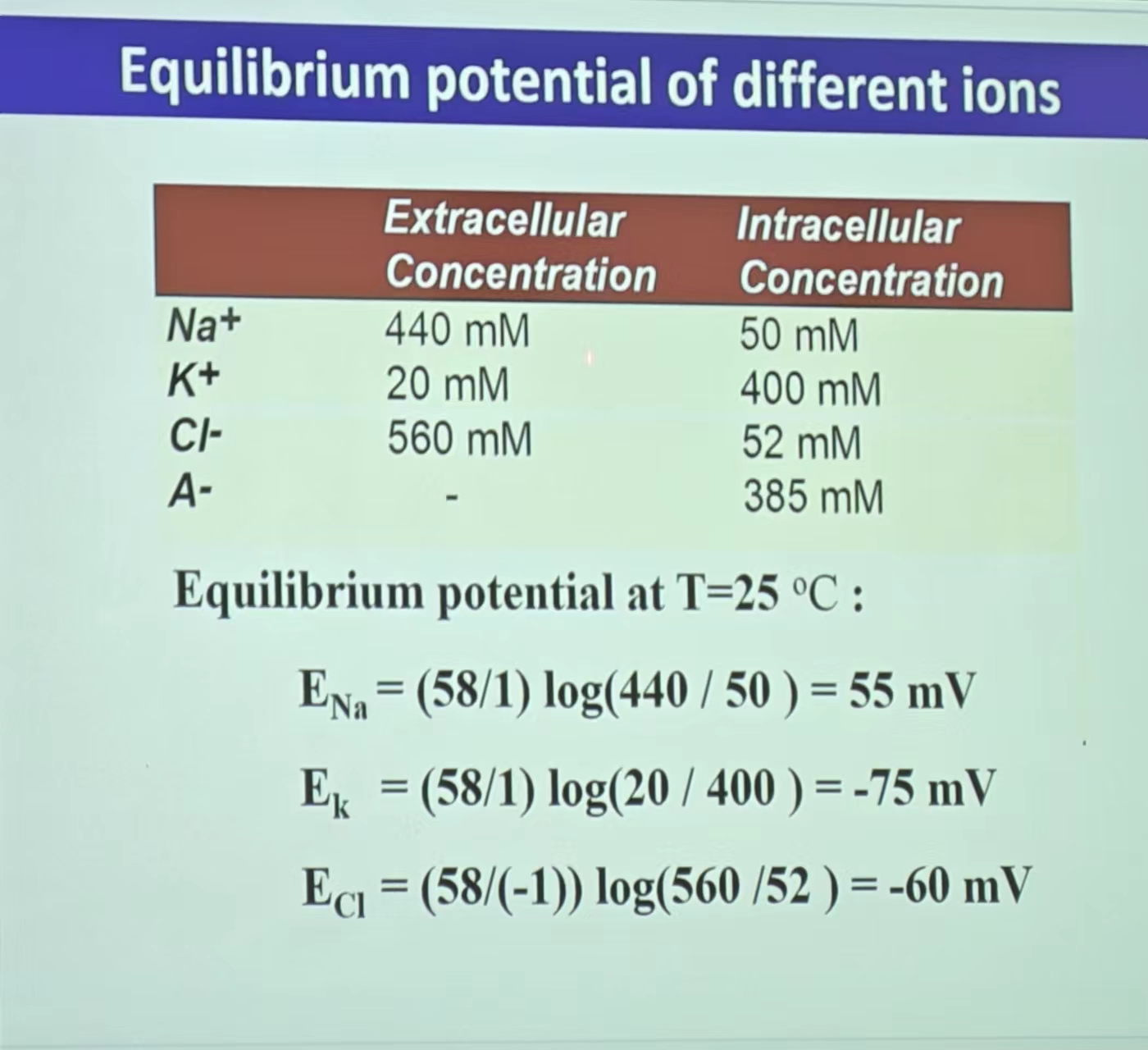
 注:在发育的不同阶段,平衡电位可能会逆转(细胞内外离子浓度的变化导致)——抑制性神经元可能向兴奋性神经元转化。
注:在发育的不同阶段,平衡电位可能会逆转(细胞内外离子浓度的变化导致)——抑制性神经元可能向兴奋性神经元转化。
- 离子平衡电位例题:
- 离子通道简化的电环路:E(单个离子通道便是类似于一个电池)
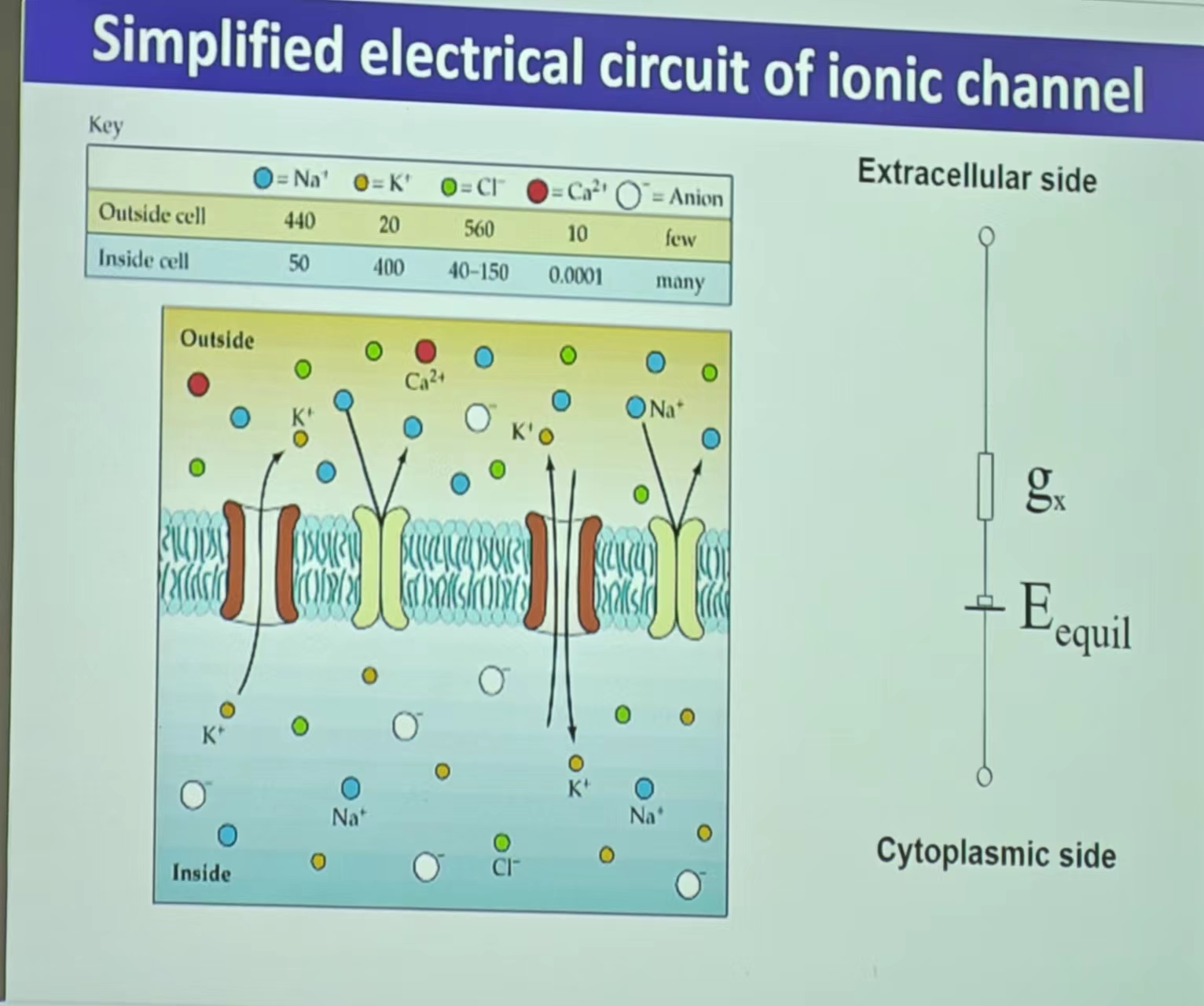
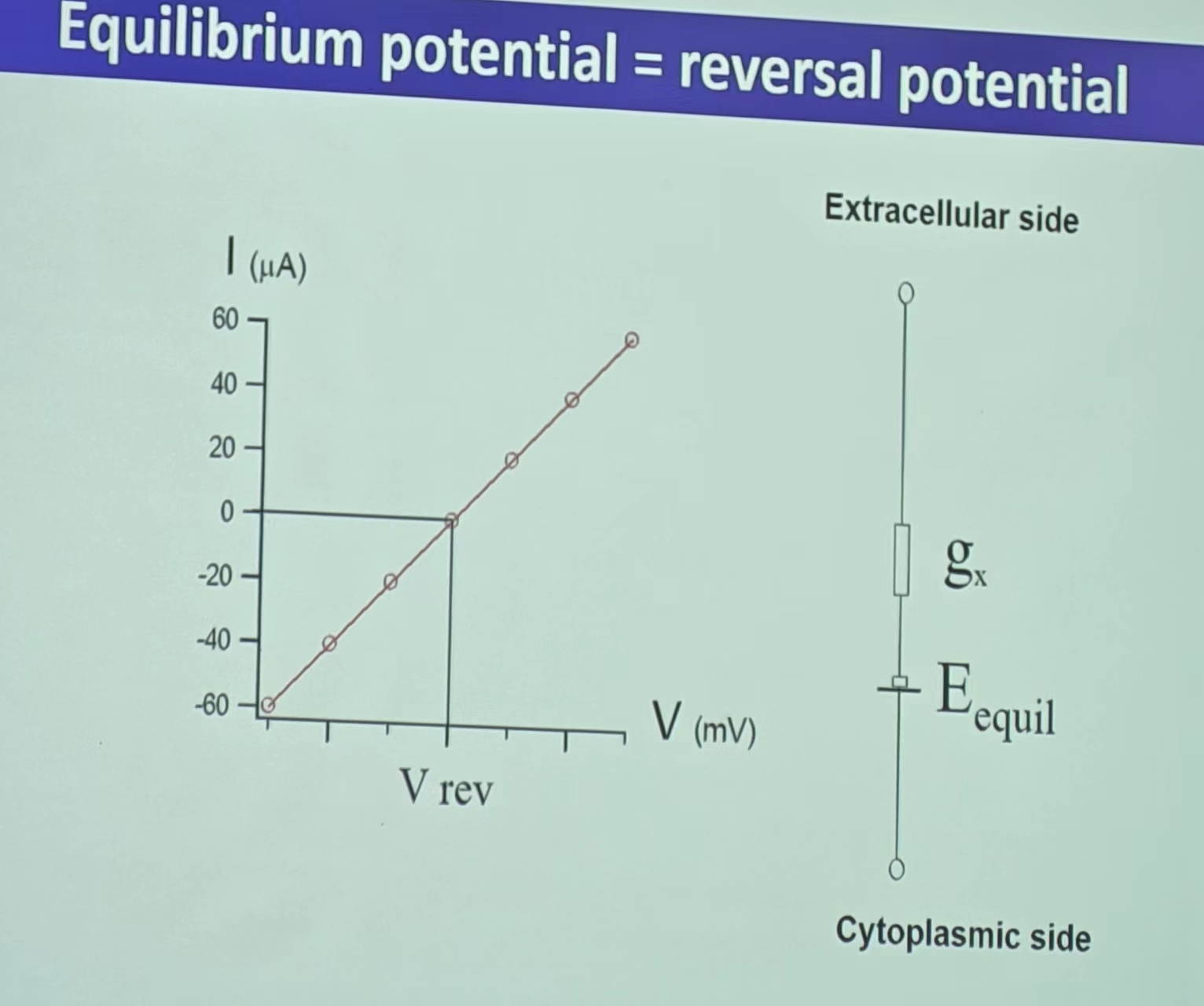 注:平衡电位又叫做翻转电位(Reversal potential),高于或者低于平衡电位的影响具体见离子运动这一部分
注:平衡电位又叫做翻转电位(Reversal potential),高于或者低于平衡电位的影响具体见离子运动这一部分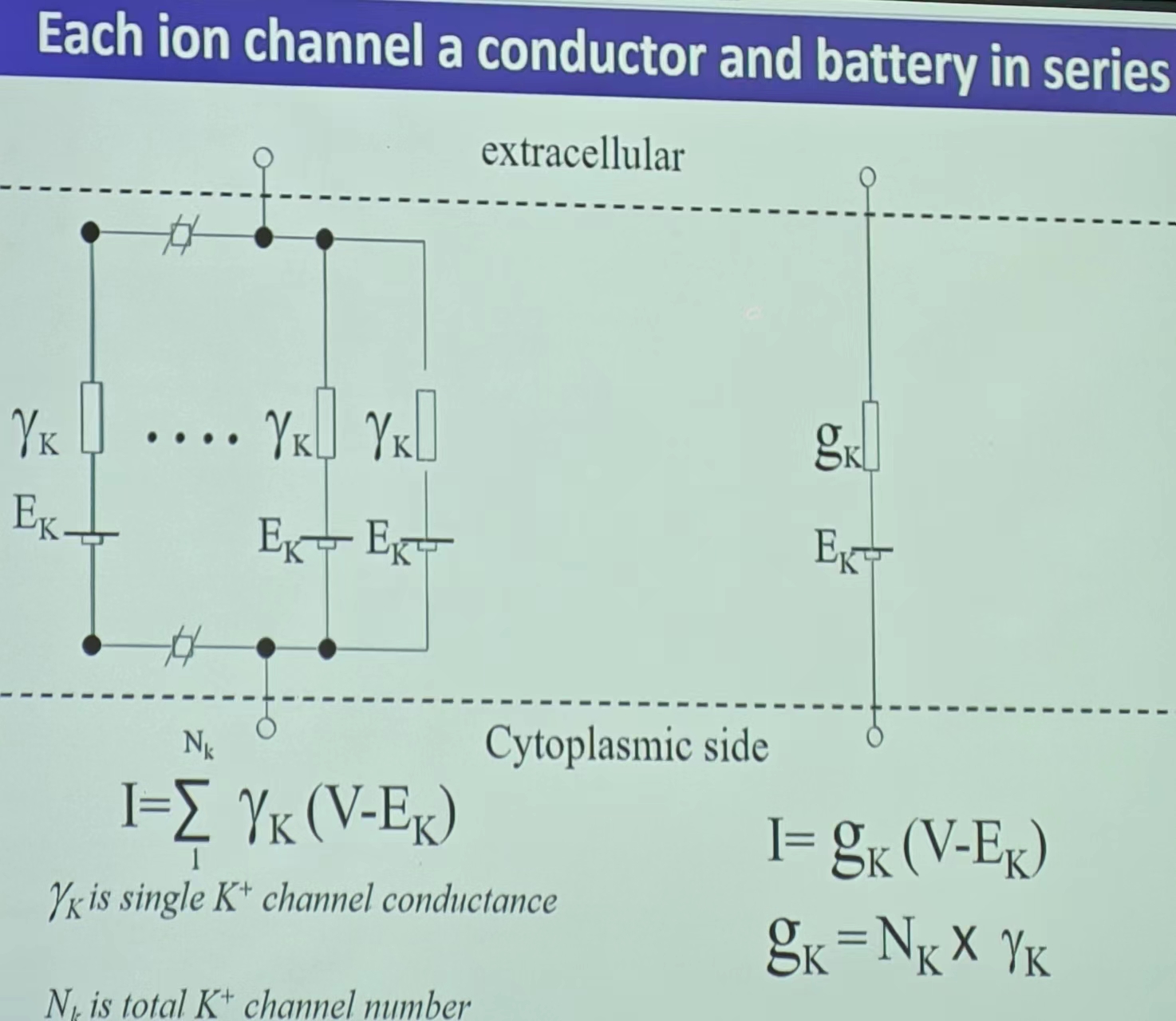
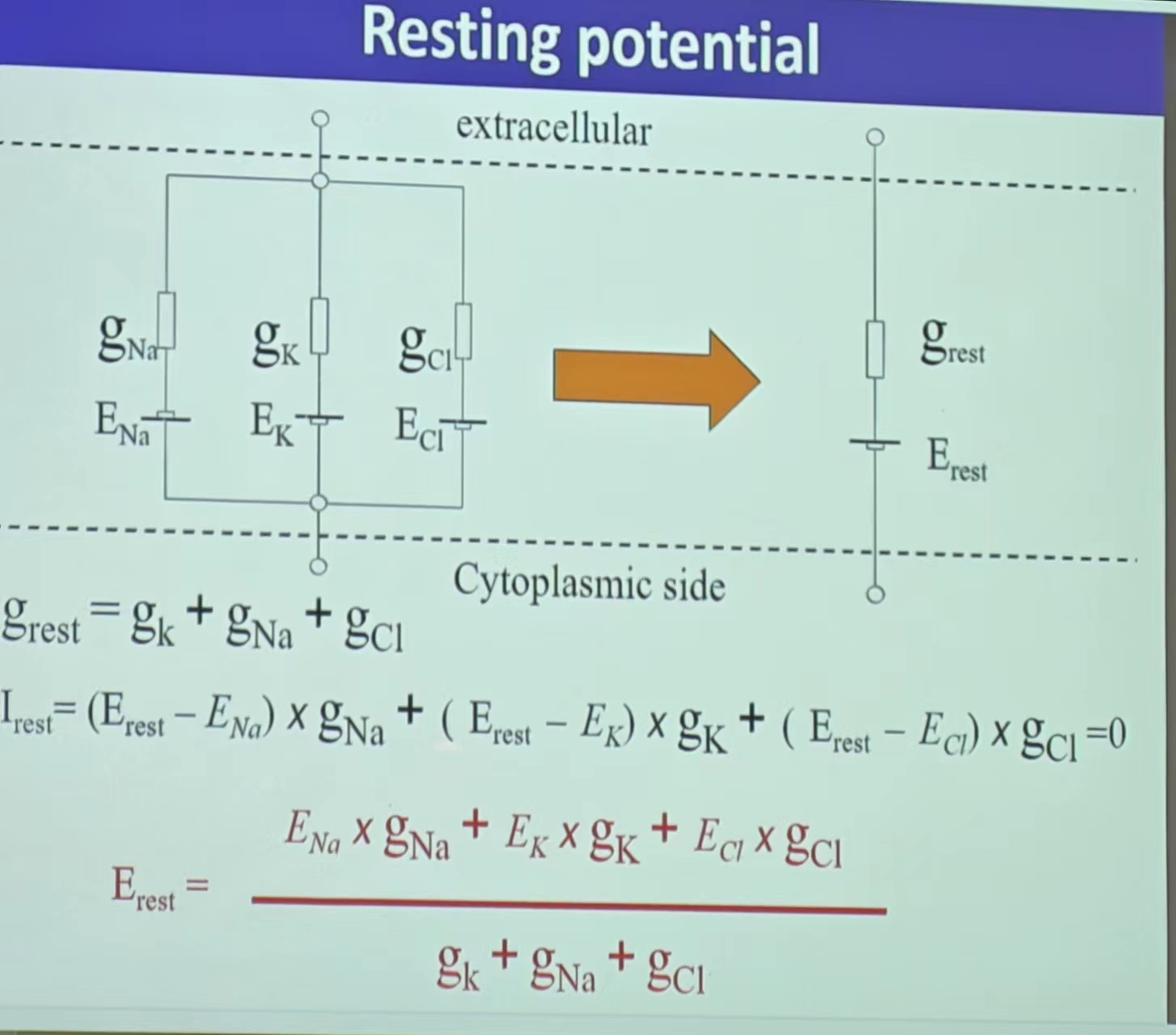
- 静息电位:INa+IK+ICl=0,即所有离子浓度处于动态平衡的状态(Na/K/Cl)——受不同离子通道的个数的影响(通透性—Probability的加权!);平衡电位导致的极化(内负外正)

-
- 离子泵:需要能量的逆流而上(防止离子一直流动的耗竭)

- Na/K泵:逆离子浓度、需ATP、可驱动其他泵(其他泵作为顺风车不一定需要ATP)

- Na/K泵:逆离子浓度、需ATP、可驱动其他泵(其他泵作为顺风车不一定需要ATP)
- 细胞电阻:受离子通道密度(越大,电阻越小)和细胞大小(越大,越小)影响
- 膜电容:此外细胞膜还是一个电容(单位面积的膜电容与膜面积一起可起算总的膜电容),细胞膜越大,膜电容越大——充电时短路,充满电断路

.jpg)
- 离子通道(电学角度:楞次电压【电压参考细胞外相对于细胞内的电压】串联电阻):
- 动作电位(firing):不符合欧姆定律的一个主动的过程(电导急剧上升,Na离子通道打开的很多),这是神经细胞的兴奋性特征(神经细胞为什么那么”神”?)
- 膜片钳:稳定电压(补偿电流),记录电流的变化——发现动作电位
- Na离子假说:A.H.—海水浓度的不同看动作电位的变化;钳住电压稳定在Na离子的平衡电位,此时I=0——A.H.提出离子通道存在的可能性
- 动作电位的产生:Na、K离子(通道是电压门控的)通透性的变化——可以通过阻断钾离子通道(4-AP)使得动作电位变宽、加河豚毒素抑制动作电位的产生
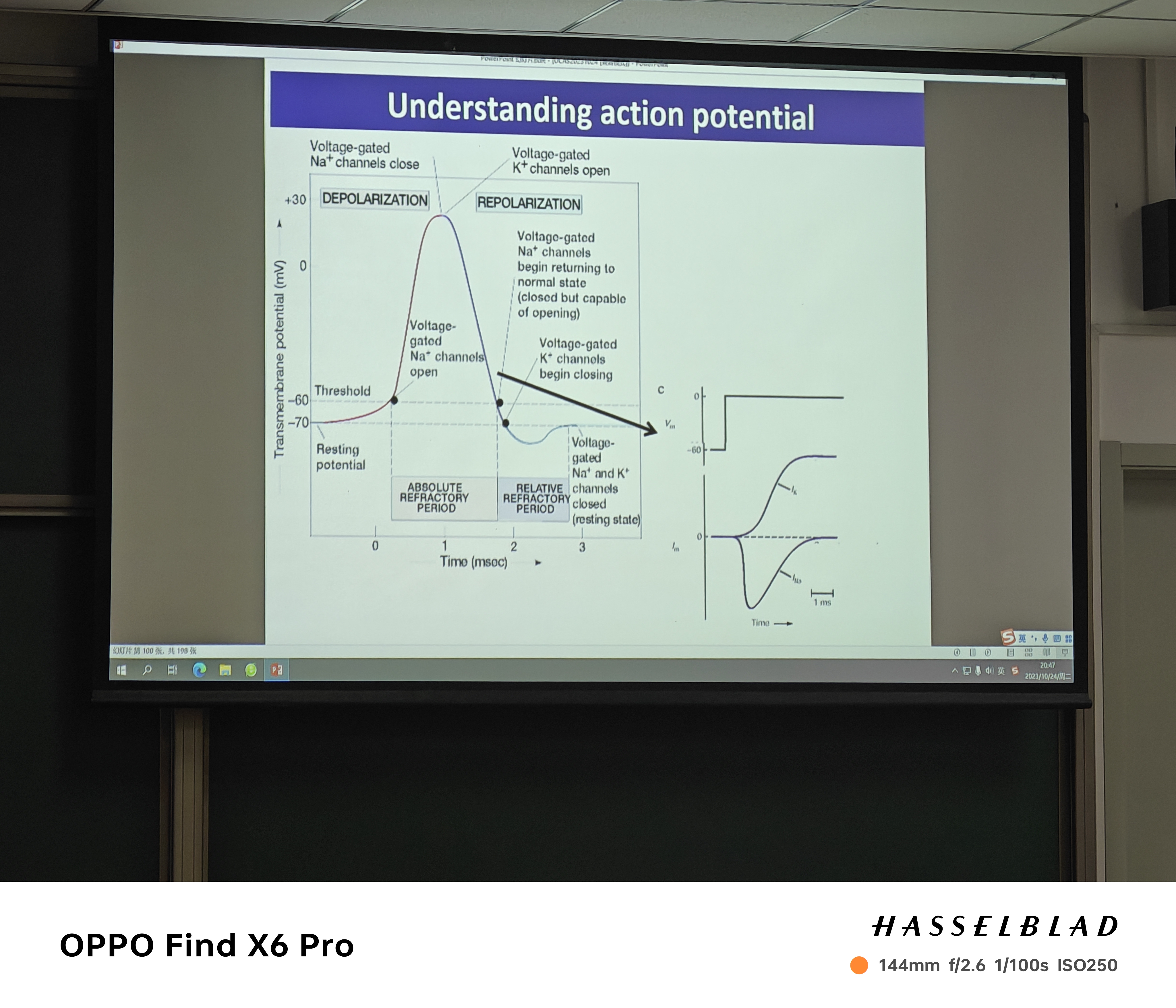
- 绝对不应期:Na离子通道失活,不能打开更多的Na离子通道
- 相对不应期:K离子通道需要打开(比静息状态打开的更多——所以要低于静息电位),K离子外流,电压下降(最后接近K离子平衡电位),需要更多的正电荷刺激才能开启动作电位。
- 注:胶质细胞、一些外周sensor不产生动作电位,但是产生去极化
- 产生过程:产生的地点究竟是什么?(追究方法:记录单位面积Na离子通道的密度+记录胞体轴突电信号产生的快慢)
- 轴突起始端产生(离开胞体50-70um)——阈值低、Na离子密度高
- 兴奋传导:沿轴突(长:存在轴向电阻、细胞膜:纵向电阻)传导;电压传导的过程中呈现指数型向刺激两端下降(decay)
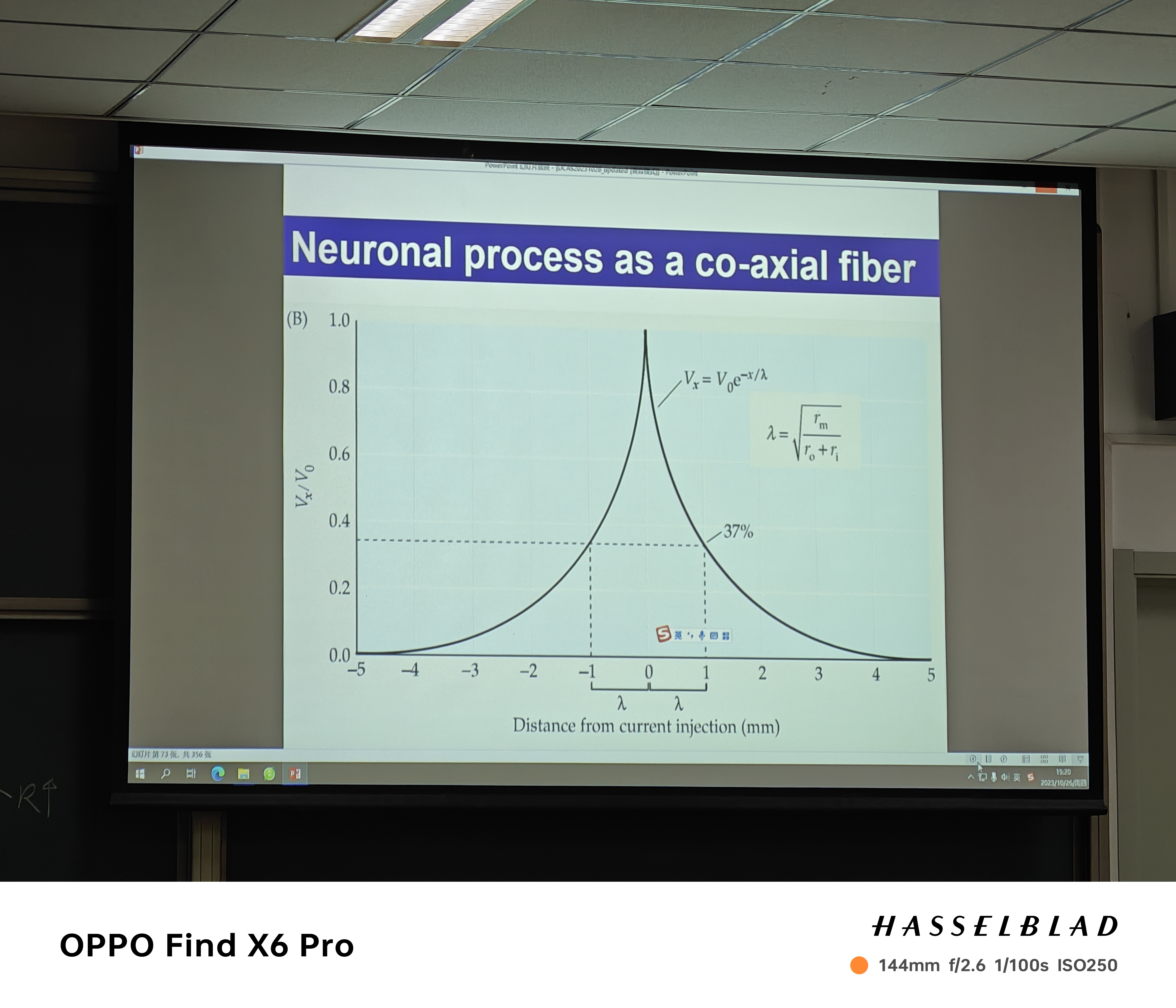
- 髓鞘:长度常数越大,电压衰减越慢,髓鞘导致长度常数变大(decay变慢);rising(charging)更快(time constent)!
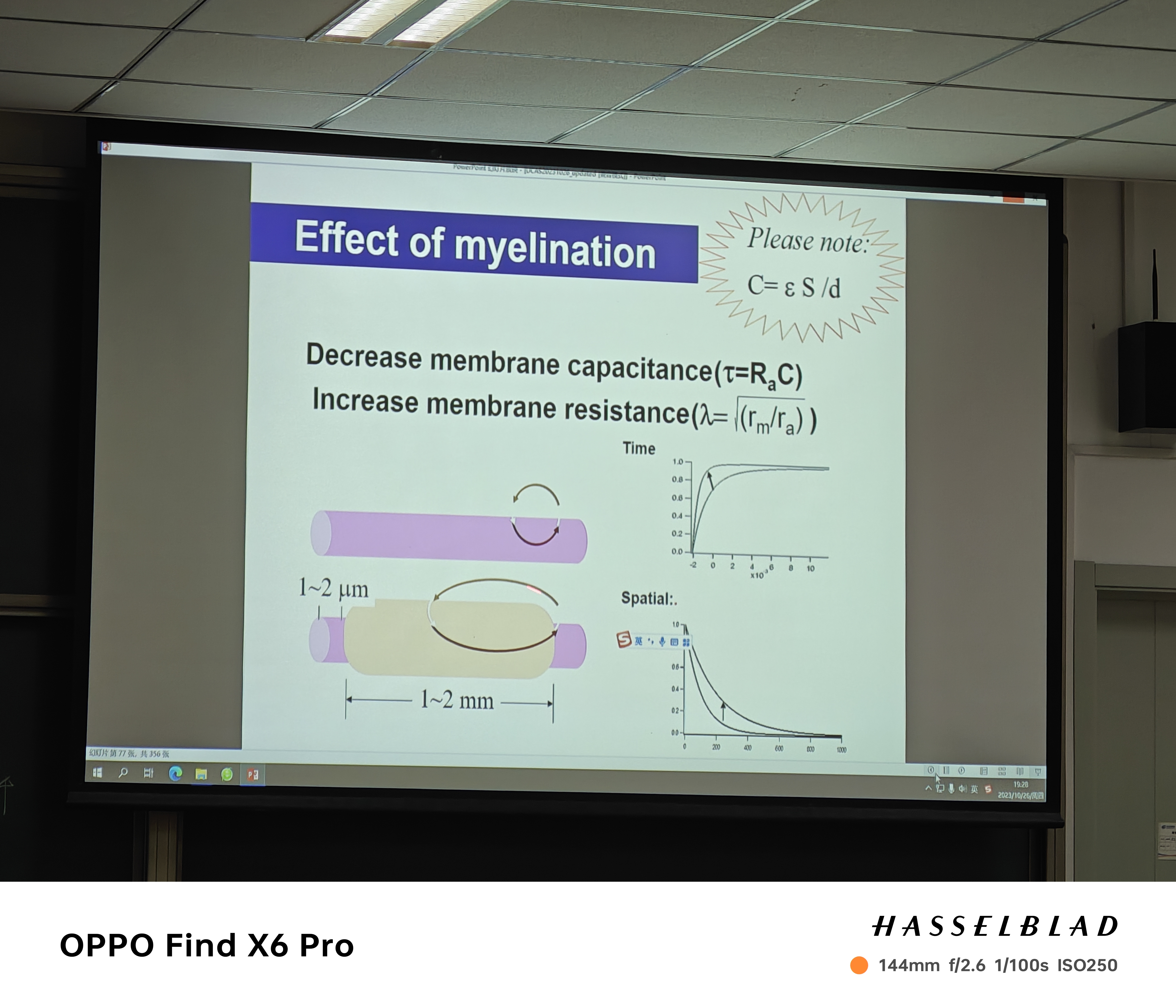
- 郎飞氏结:跳跃式传导(髓鞘存在的结果)
- 髓鞘:长度常数越大,电压衰减越慢,髓鞘导致长度常数变大(decay变慢);rising(charging)更快(time constent)!
- 输入:其他神经细胞通过突触兴奋下游神经元
- 突触(神经信息处理的最小单位):
- 突触的重要性在哪?:例:NR2B过表达——老鼠变聪明
- Concept(几百nm级别):
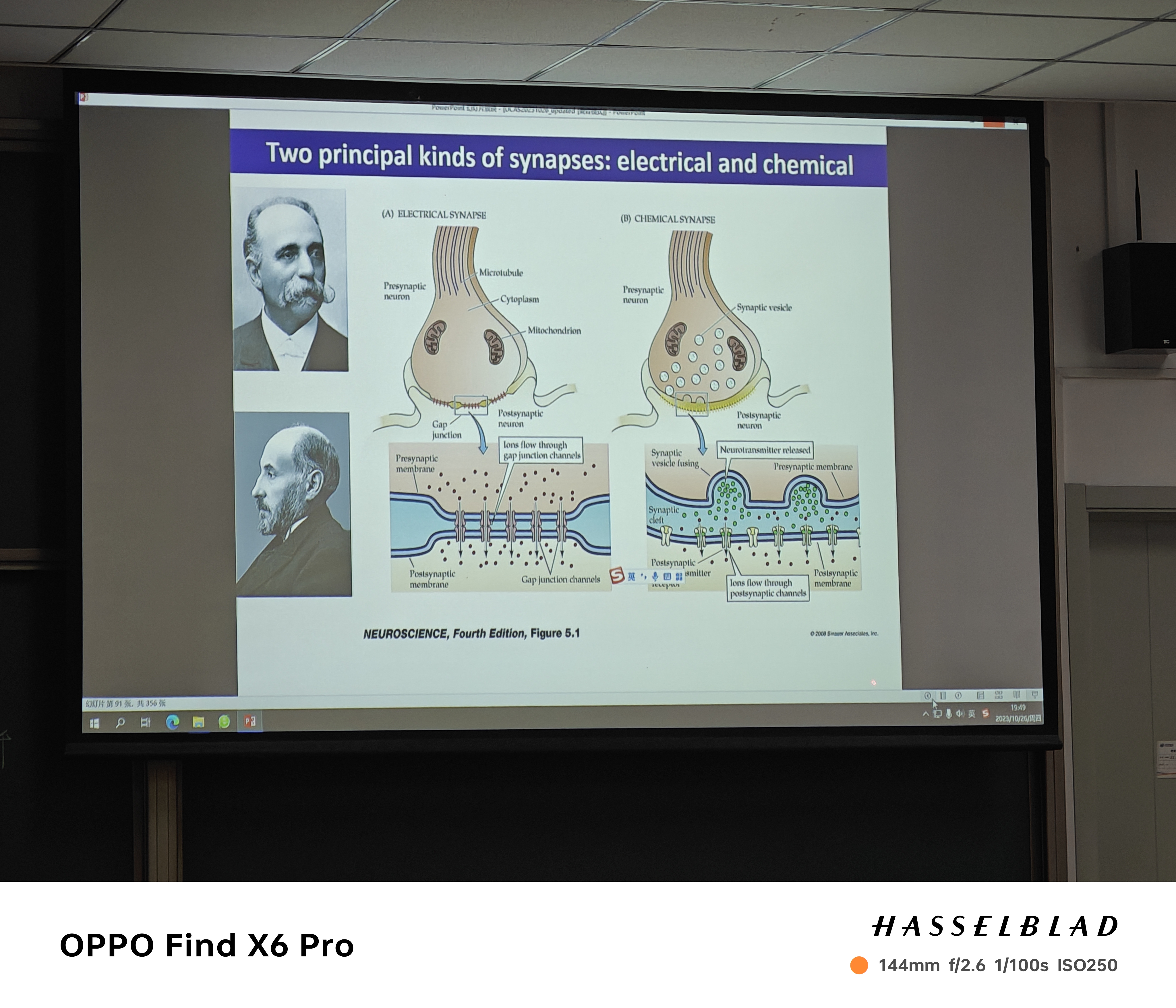
- 化学突触(卡哈尔):分离的状态(迷走神经刺激心脏的过程),存在delay——中枢神经系统90%为化学突触
- 含轴突-轴突、轴突-胞体、轴突-树突的连接方式
 注:非对称的前膜和后膜介导兴奋,对称介导抑制
注:非对称的前膜和后膜介导兴奋,对称介导抑制 - 神经肌肉接头NMJ也是化学突触
- 含轴突-轴突、轴突-胞体、轴突-树突的连接方式
- 电突触(Golgi):传导速度非常快,小于1ms——需要同步活动的神经细胞存在(gap junction)——上游或者下游的细胞死亡后一定一直关闭,PH过高也会导致关闭。
- coupling coefficient:下游的反应/上游的反应(K ratio-谁大谁当分子,即为上游)

- 树突不是绝对的输入点(树突对树突)
- connexons为六聚体,六个connexin组成

- coupling coefficient:下游的反应/上游的反应(K ratio-谁大谁当分子,即为上游)
- 化学突触(卡哈尔):分离的状态(迷走神经刺激心脏的过程),存在delay——中枢神经系统90%为化学突触
- 突触传递:
- 神经肌肉接头上的发现:Katz——钙离子依赖的mEPP的发现(自发的noise),即神经信号的最小单位(quanter)——囊泡和量子化神经递质释放导致的结果(囊泡释放的独立性—满足二项分布)
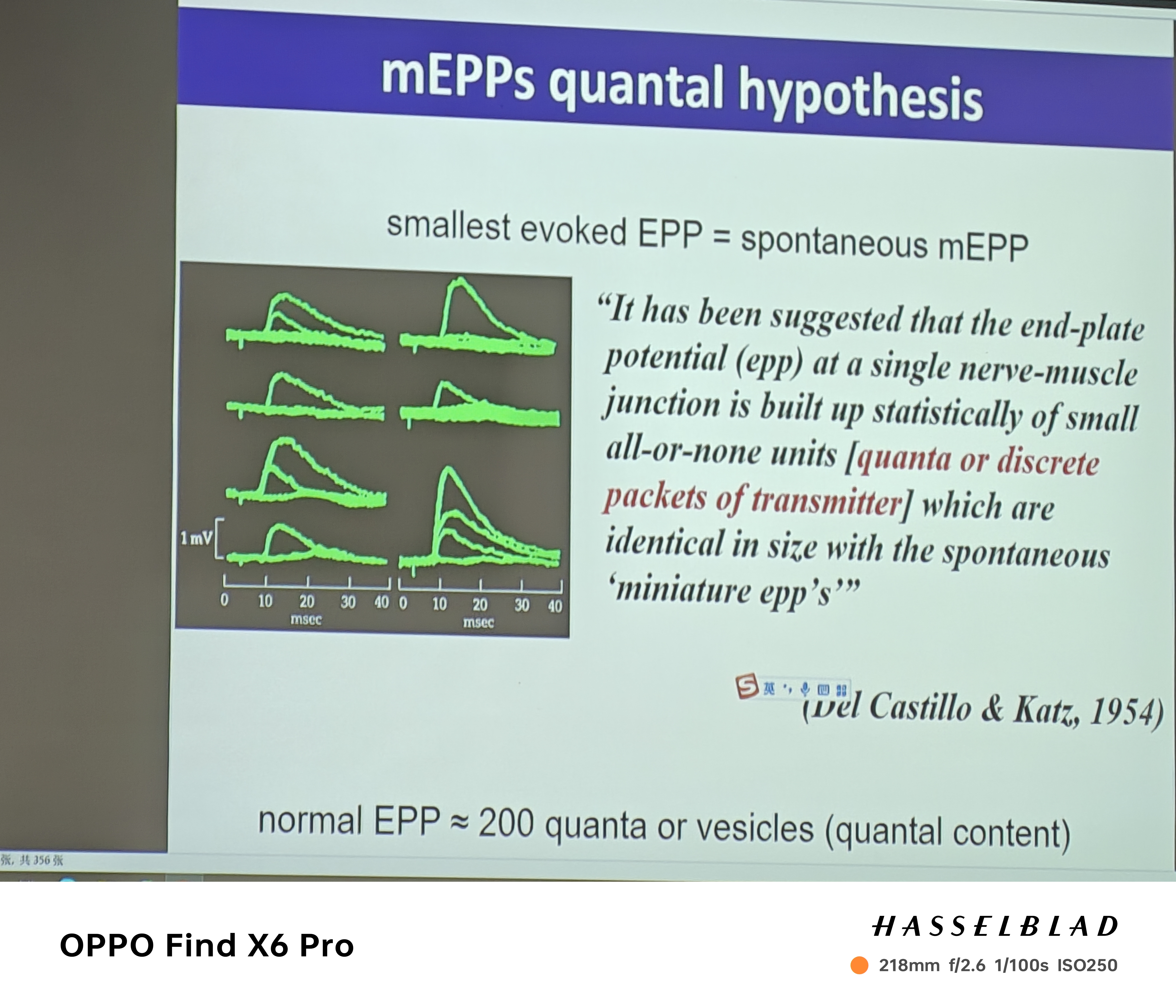
- 传递过程(动作电位来时,囊泡以一定可能性释放):
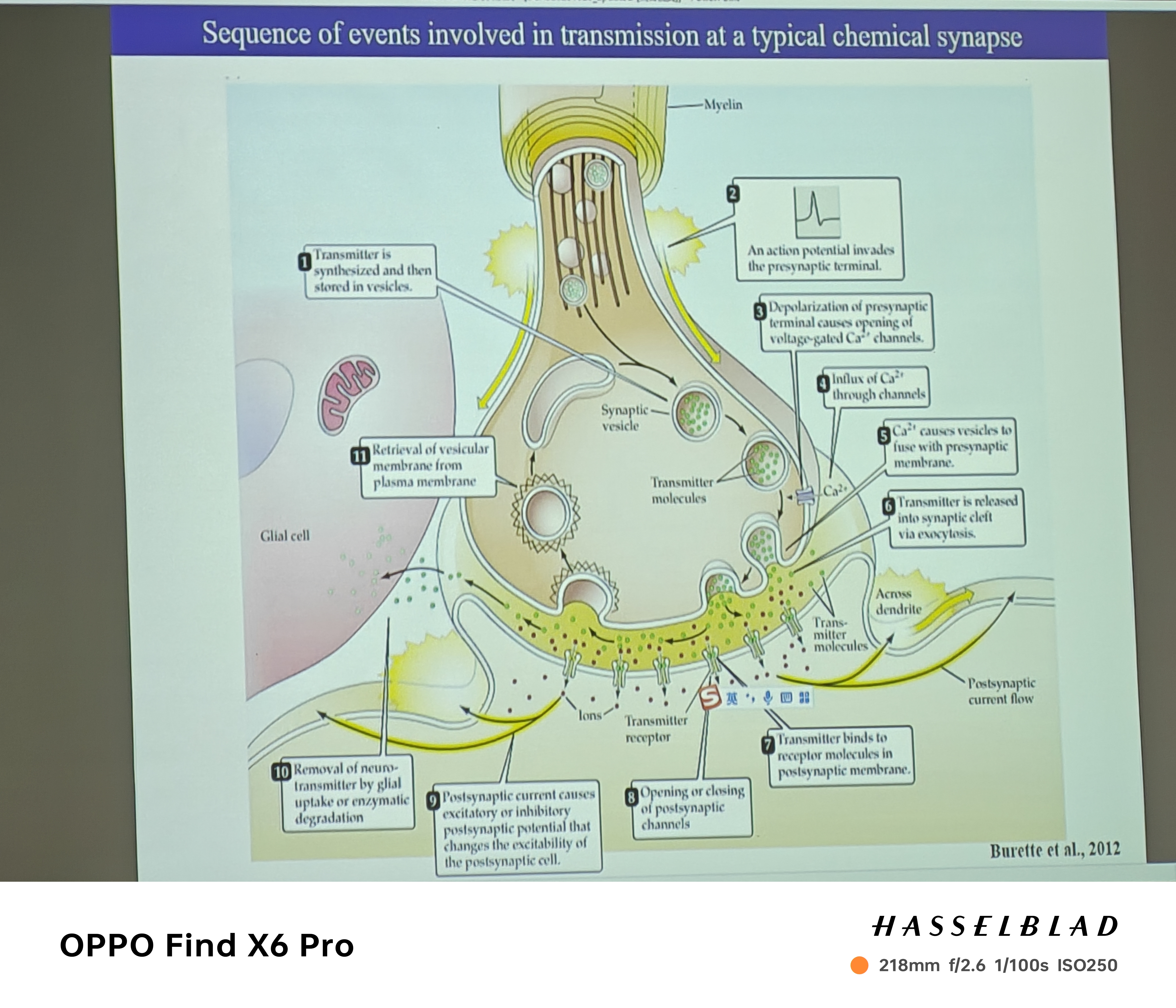
- 膜电容的突然变大(囊泡的融合的过程,即神经递质的释放导致细胞膜表面积变大)
- 神经递质的释放就是膜融合的过程吗?——4AP导致神经递质释放的更多,同时膜电容也变的越大,因此是同时的

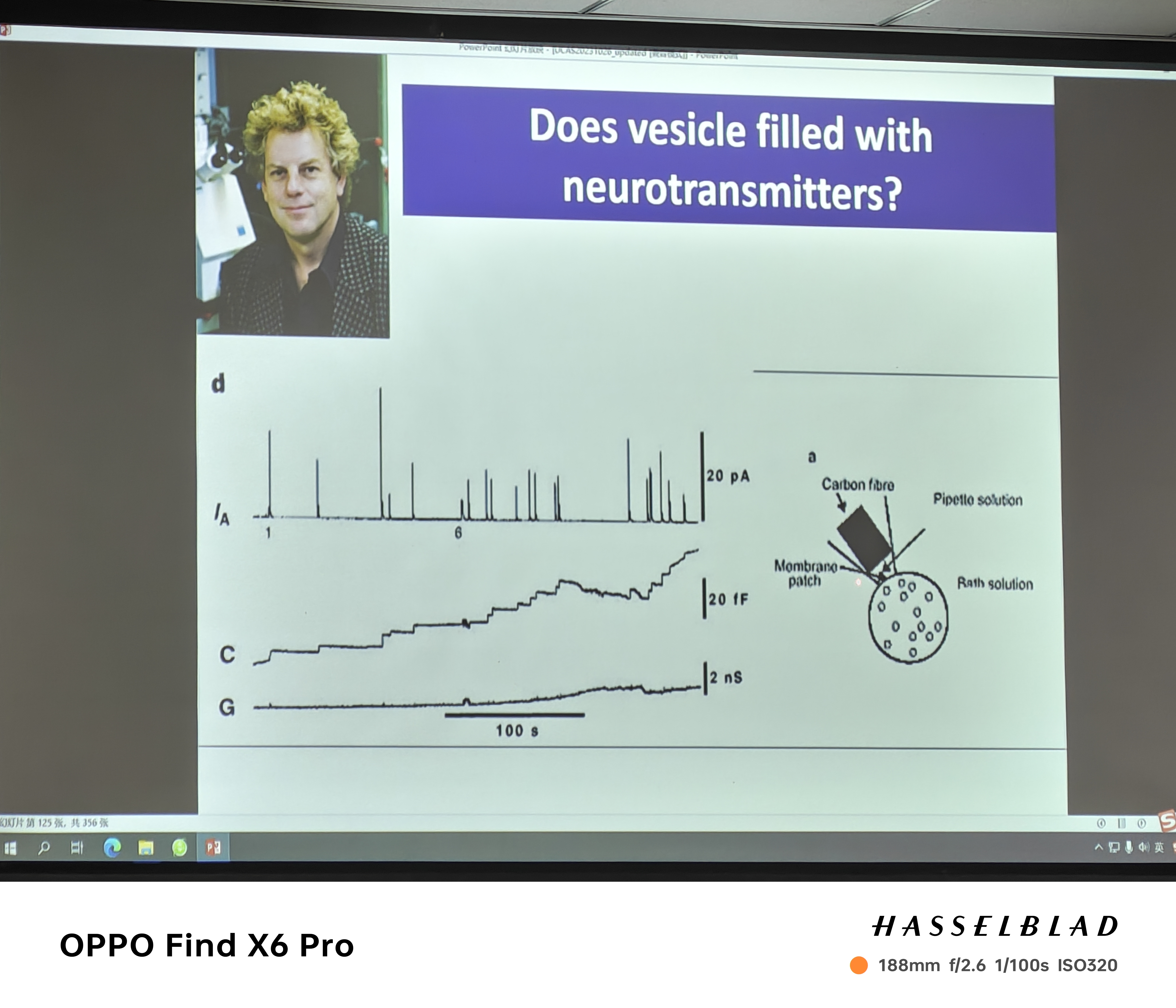
- Ca离子通道与囊泡耦合吗?——(1)囊泡(synaptotagmin)和钙离子通道的蛋白分别标记荧光,看看是否Merge (2)氯化镉阻断钙离子通道——膜电容不变 (3)BAPTA:钙离子缓冲剂(半道拦截钙离子),钙离子内流,但膜电容几乎不变——非钙离子通道打开才导致囊泡释放 (4)钙离子螯合剂(DM)——紫外线导致uncage,所有钙离子都释放出,突触末端所有位置钙离子浓度都一致,控制了单一变量——记录光活化后膜电容的变化情况
- 神经递质的释放就是膜融合的过程吗?——4AP导致神经递质释放的更多,同时膜电容也变的越大,因此是同时的
- 囊泡结合到active zone后发生内吞——膜电容的drop
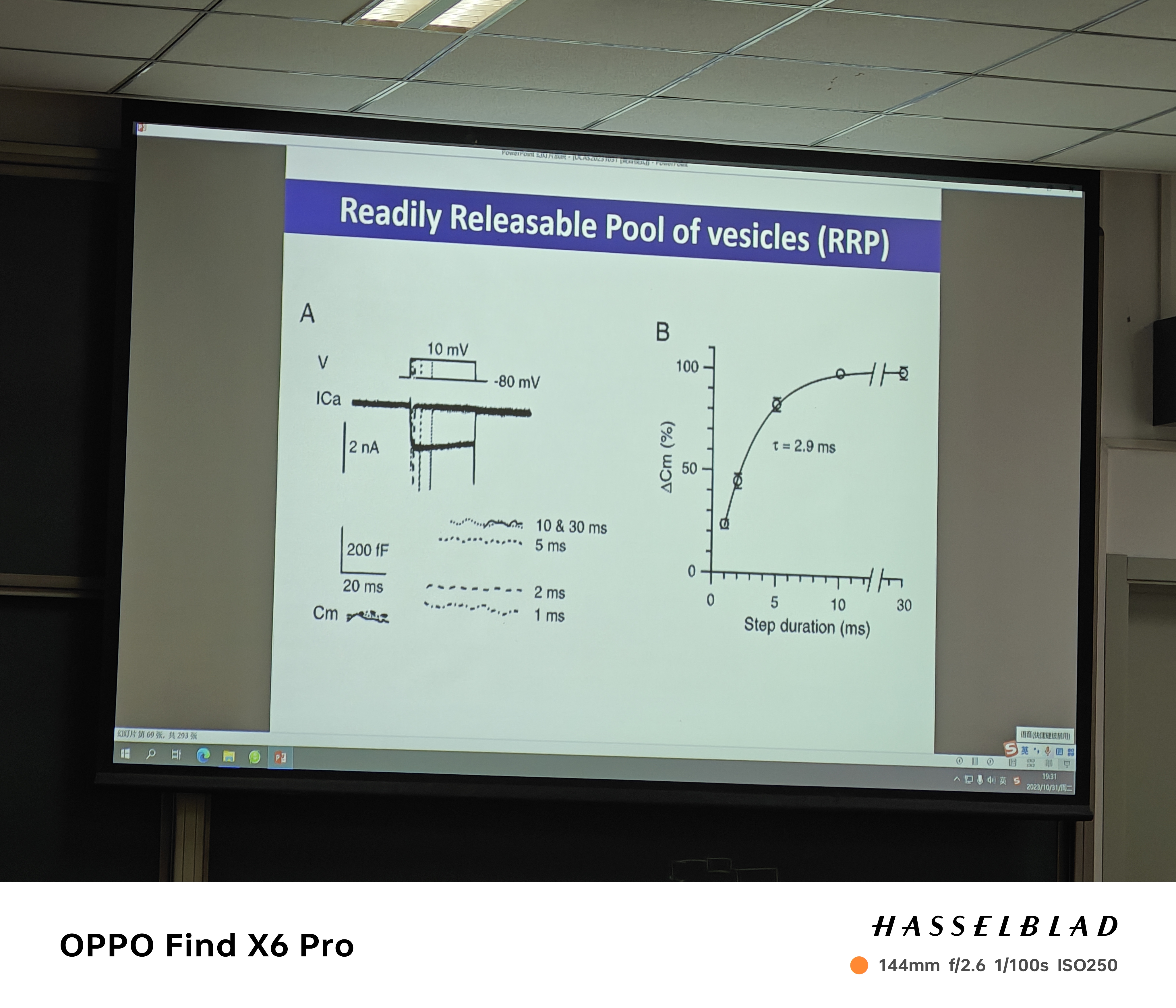
- RRP(readily releasable pool of vesicles):随着刺激的增加,囊泡释放的数量增加,然后数目增加不动,膜电容最后不变
 注:其中最大的一群由synapsin维持在突触前膜远端,叫做储备池(Reserve Pool),它们只有被定位到活性区成为即刻可释放池(Readily Releasable Pool, RRP)才有机会被释放,另有一小亚群在两者之间穿梭,又称循环池(Recycling Pool)
注:其中最大的一群由synapsin维持在突触前膜远端,叫做储备池(Reserve Pool),它们只有被定位到活性区成为即刻可释放池(Readily Releasable Pool, RRP)才有机会被释放,另有一小亚群在两者之间穿梭,又称循环池(Recycling Pool)
- 有多少囊泡释放呢?——EPSC见上图
- priming(囊泡的预备和成熟):填充囊泡的过程——可记录零点即开始时有多少囊泡(黑线是replenishment——预测曲线,预测RRP,红线是囊泡释放数量(囊泡融合的过程),零点时,R-P代表的就是RRP)
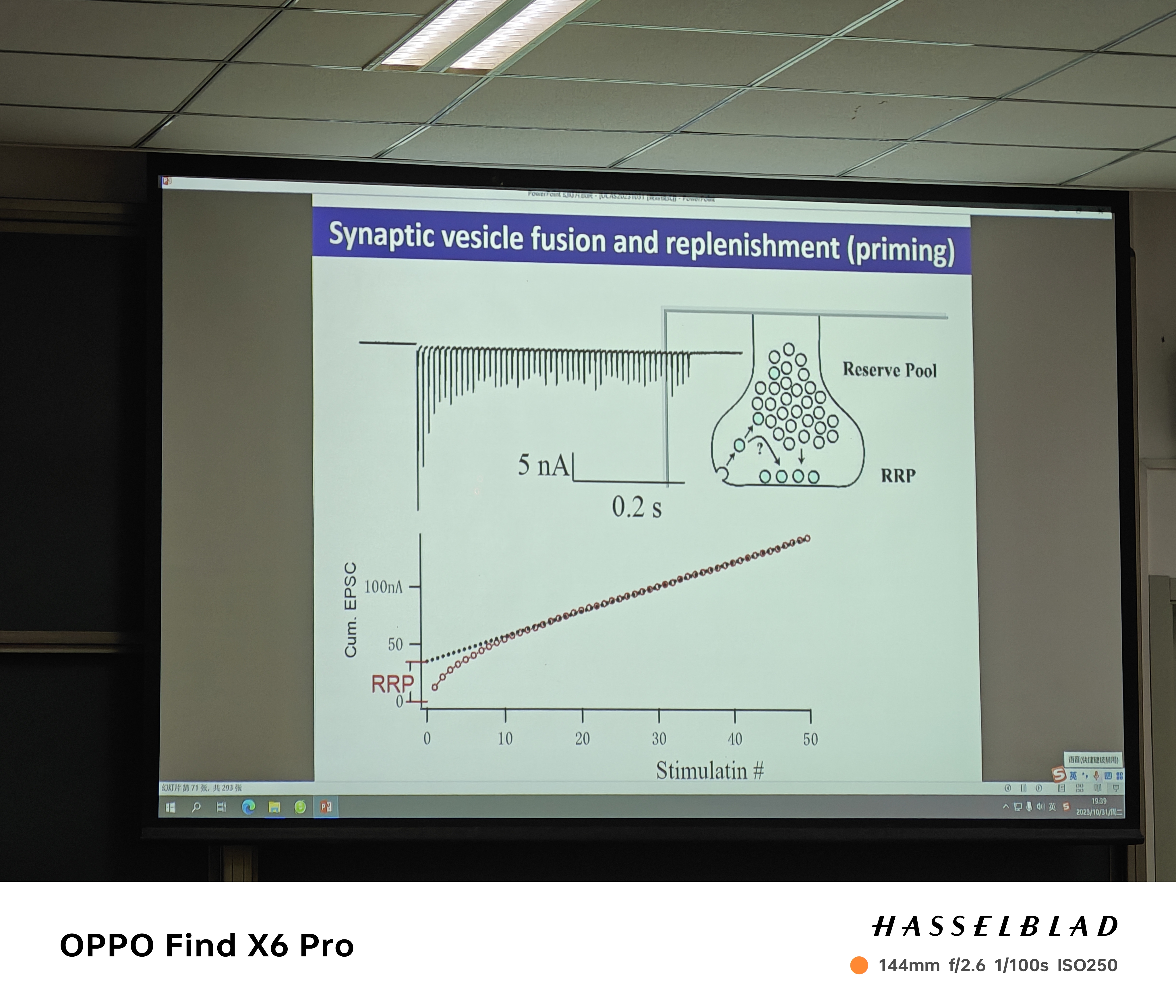 注:随着刺激的增加,EPSP降低然后保持不变——RRP数量有限,Reserve pool释放需要时间且缓慢(释放和回收的过程需要一定的时间,在下一次刺激的适合仅有少量囊泡可供释放,因为有一部分局限在在回收的过程中。然后随着刺激的增加,这个时间差会达到一个阈值(随着刺激增加,囊泡释放保持不变时),EPSP也保持不变了。)
注:随着刺激的增加,EPSP降低然后保持不变——RRP数量有限,Reserve pool释放需要时间且缓慢(释放和回收的过程需要一定的时间,在下一次刺激的适合仅有少量囊泡可供释放,因为有一部分局限在在回收的过程中。然后随着刺激的增加,这个时间差会达到一个阈值(随着刺激增加,囊泡释放保持不变时),EPSP也保持不变了。) - 囊泡回收的酸化过程?(刺激的时候PH中性化)——利用PH-senstive的荧光标记囊泡蛋白(回收时酸化导致荧光变亮?)。若要避免漂白(NH4Cl的加入导致荧光上升,则不是Bleaching)

- 囊泡的循环:FM-dye(刺激变亮(吸收dye),wash再刺激变暗说明循环)

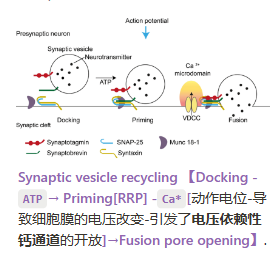
- 膜电容的突然变大(囊泡的融合的过程,即神经递质的释放导致细胞膜表面积变大)
- 神经肌肉接头上的发现:Katz——钙离子依赖的mEPP的发现(自发的noise),即神经信号的最小单位(quanter)——囊泡和量子化神经递质释放导致的结果(囊泡释放的独立性—满足二项分布)
- 囊泡融合和释放的机制(脂质体的融合)
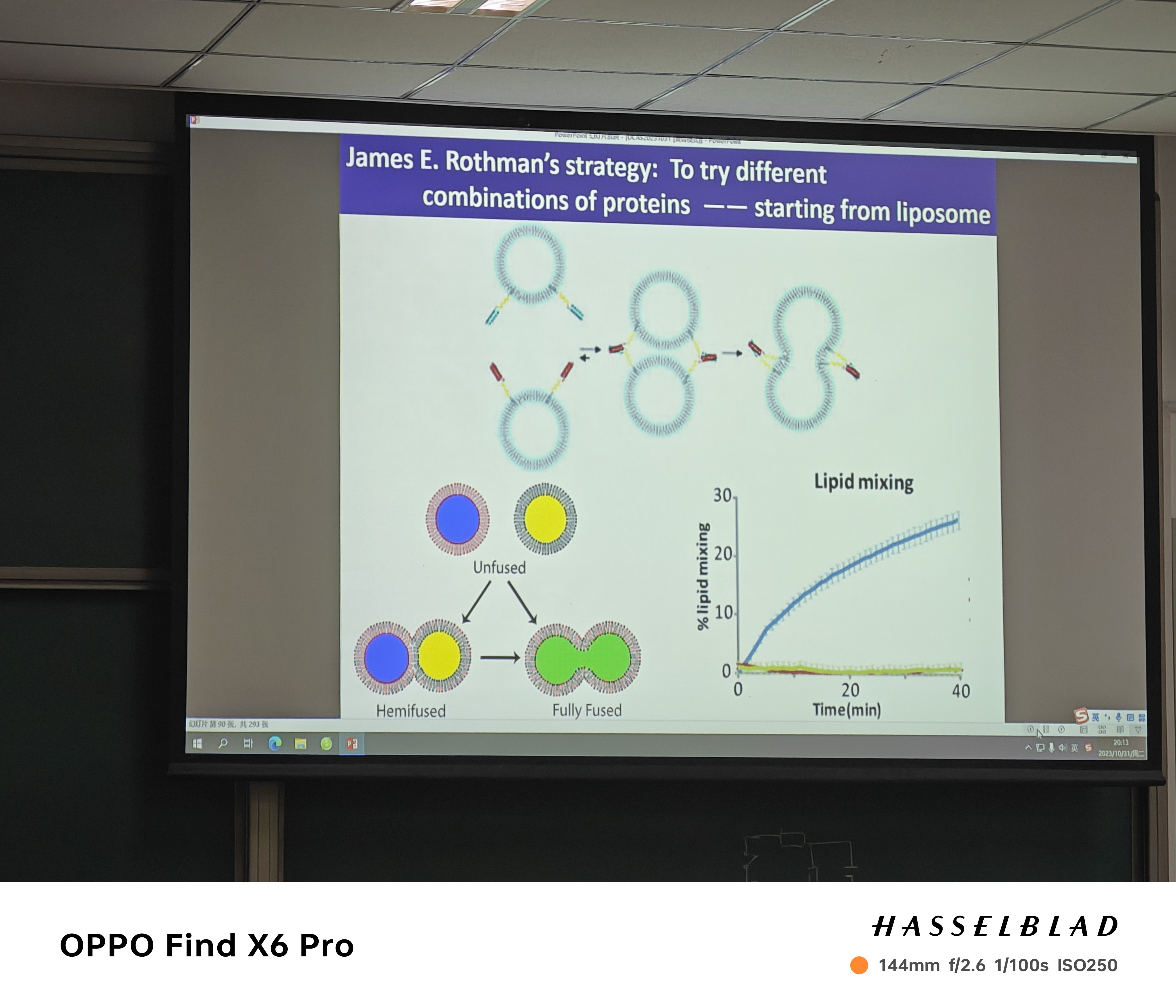
- 1.发现:Randy——酵母突变体的筛选(fusion难以work);James——尝试再两个囊泡表达不同蛋白(带不同颜色),融合即变色——融合说明两蛋白存在融合机制
-
- 融合机制:SNARE-complex发挥的功能(诺奖),这是充分必要条件


- 融合机制:SNARE-complex发挥的功能(诺奖),这是充分必要条件
- 3.融合时间慢,但突触传递快?
- 快释放;Thomas——钙离子传感蛋白(囊泡上,有五个钙离子结合位点,可以与snare结合),有助于快速的突触传递(囊泡释放前)。证明——没这个蛋白,突触传递就不会很快;敲掉之后再overexpress,又可以rescue;在钙离子结合位点做Mutation,突触传递对钙离子的敏感度变弱;把affinity增加,突触传递对钙离子的敏感性增加
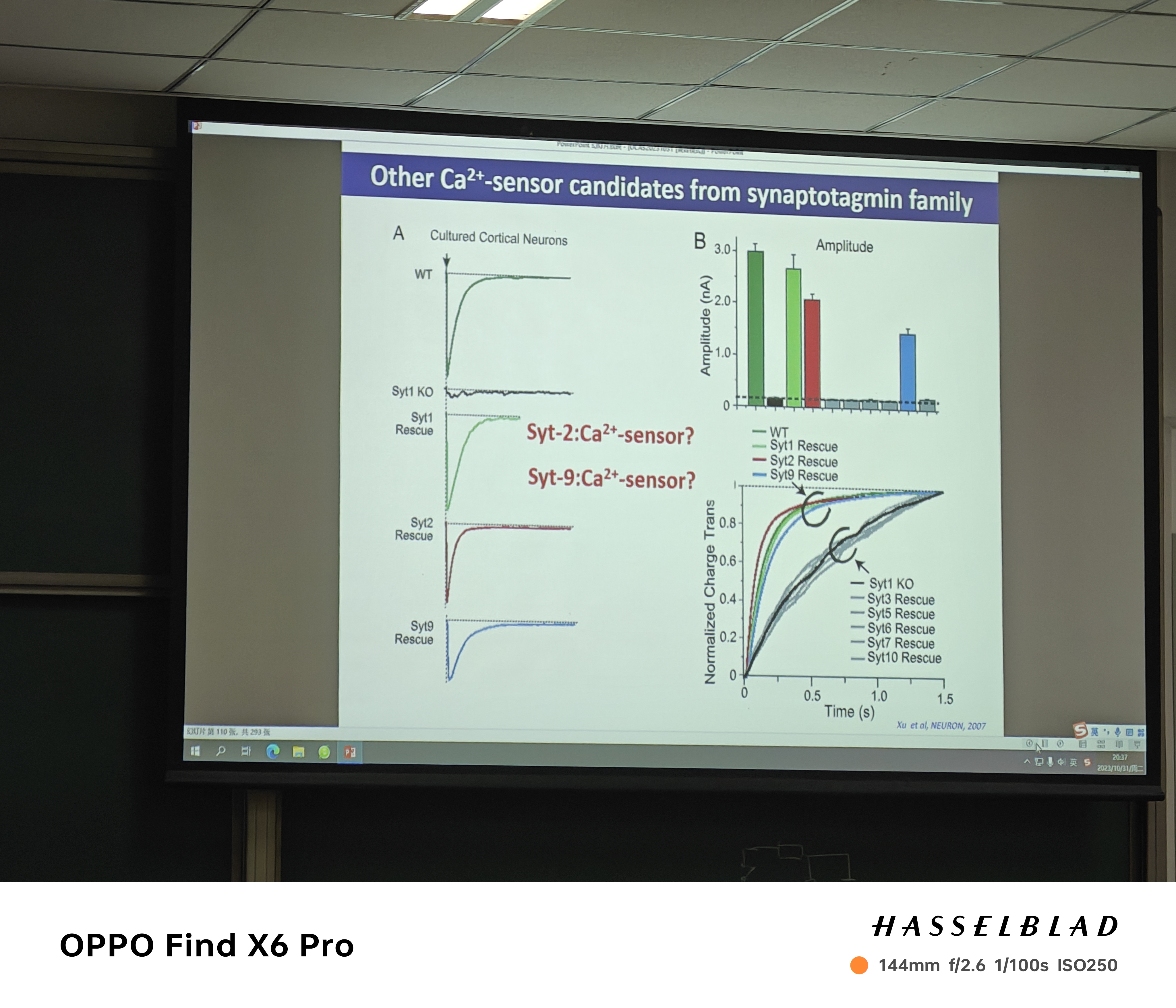
- 慢/异步释放(停止刺激,突触后膜还有动作电位)——也存在传感蛋白?
- Calyx of held synapse:syt1(囊泡上)/9没有,syt2有(对快速传递有作用)
- slope~2:两个钙离子bindsite的发现
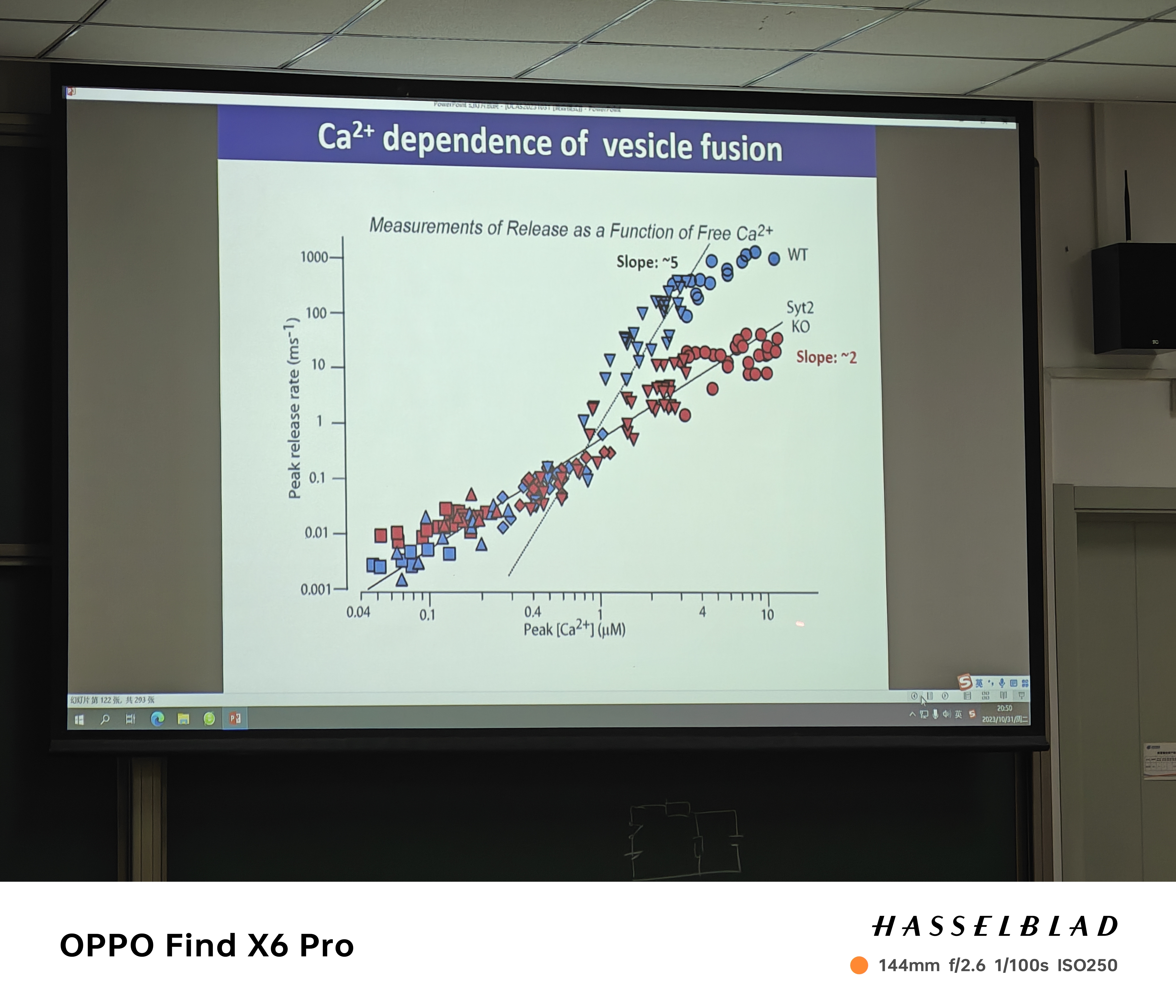
- 两个传感系统是否存在竞争?互相做stabililty的抑制,是否出现此消彼长的过程;

- 谁是slow sense?——syt7(nerve teminal细胞膜上),slow sense的存在导致囊泡的非同步释放

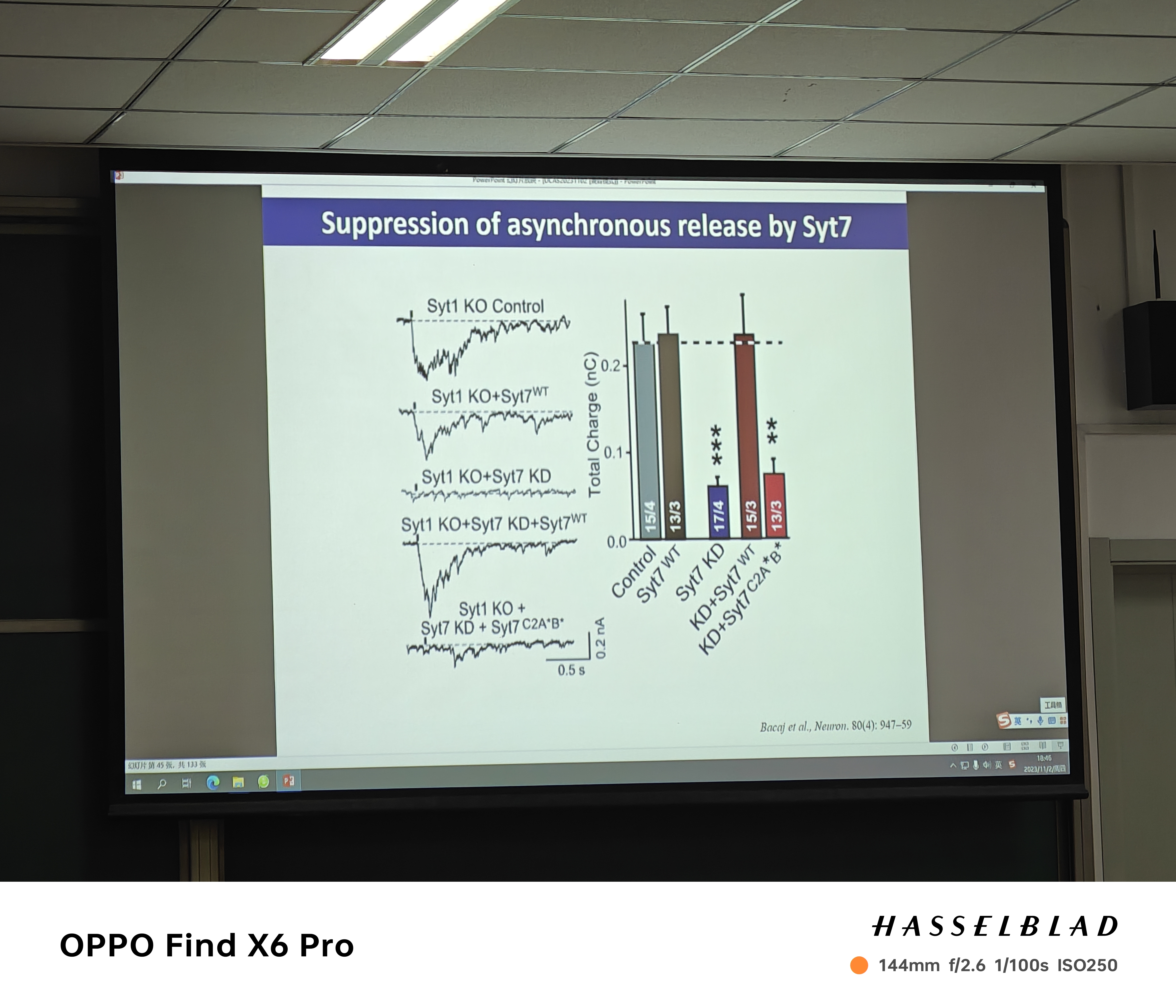
- 快释放;Thomas——钙离子传感蛋白(囊泡上,有五个钙离子结合位点,可以与snare结合),有助于快速的突触传递(囊泡释放前)。证明——没这个蛋白,突触传递就不会很快;敲掉之后再overexpress,又可以rescue;在钙离子结合位点做Mutation,突触传递对钙离子的敏感度变弱;把affinity增加,突触传递对钙离子的敏感性增加
- 1.发现:Randy——酵母突变体的筛选(fusion难以work);James——尝试再两个囊泡表达不同蛋白(带不同颜色),融合即变色——融合说明两蛋白存在融合机制
- 囊泡的内吞:Clathrin的组装(有助于内吞体形成新的囊泡)、Dynamin(actin)的作用(GTP的水解提供能量)

- 神经递质(调质):


- 判断条件:在突触前合成、Ga离子驱动释放的、突触后需要受体
- Pathways:可见弥散神经系统
- Cholinergic(胆碱能):由丘脑的胞体投射
- 多巴胺能:黑质
- 去甲肾上腺素:蓝斑、外侧被盖区
- 5-HT:中缝核
- VTA-NAc:奖赏环路
- 神经递质受体(配体耦合):

- 离子通道型受体
- 如何检测离子通道通过的离子?——改变细胞外特定离子的浓度,看电流的变化情况
- 兴奋or抑制?:与reversal potential(平衡电位)决定(Nernst方程)——发育的早期,Cl离子细胞内浓度有所不同,这时候它的平衡电位不同(起兴奋性作用)
- G蛋白偶联受体(代谢性受体)

- 离子通道型受体
- 神经递质的循环:Ach(小分子才循环,神经肽不循环)
- EPSC的叠加效应:时间和空间的整合

- 历史脉络:(1)Galvani通过蛙的坐骨神经发现生物电
突触可塑性:Fire together, wire together
- 海马内的LTP:突触前的强刺激,直接刺激突触后神经元(皆产生LTP);突触前持续的弱刺激(LTD)
- ——大部分的LTP产生在突触后神经元(原理:AMPA受体的上膜、NMDA也在LTP中起关键作用,无NMDA则无法产生LTP——NMDA受体影响了AMPA的上膜——钙离子低于平衡电位内流导致的结果)

 注:LTP诱导先激活突触后NMDA受体,造成NMDA受体开放,胞外钙通过该通道进入胞内,相继激活下游的CaMKII、Ras、PI3K,进而磷酸化AMPA受体的GluA1亚单位,造成其活动依赖性的转运上膜。
注:LTP诱导先激活突触后NMDA受体,造成NMDA受体开放,胞外钙通过该通道进入胞内,相继激活下游的CaMKII、Ras、PI3K,进而磷酸化AMPA受体的GluA1亚单位,造成其活动依赖性的转运上膜。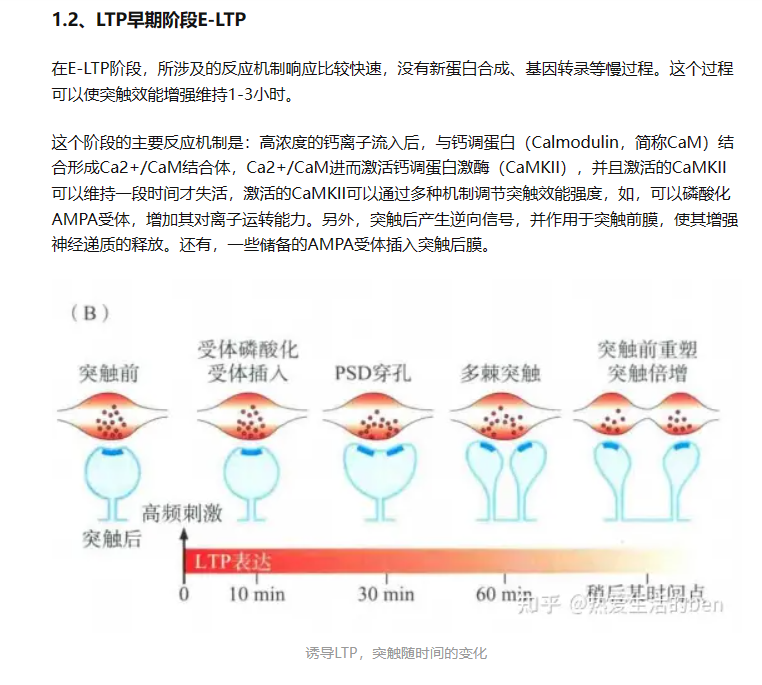

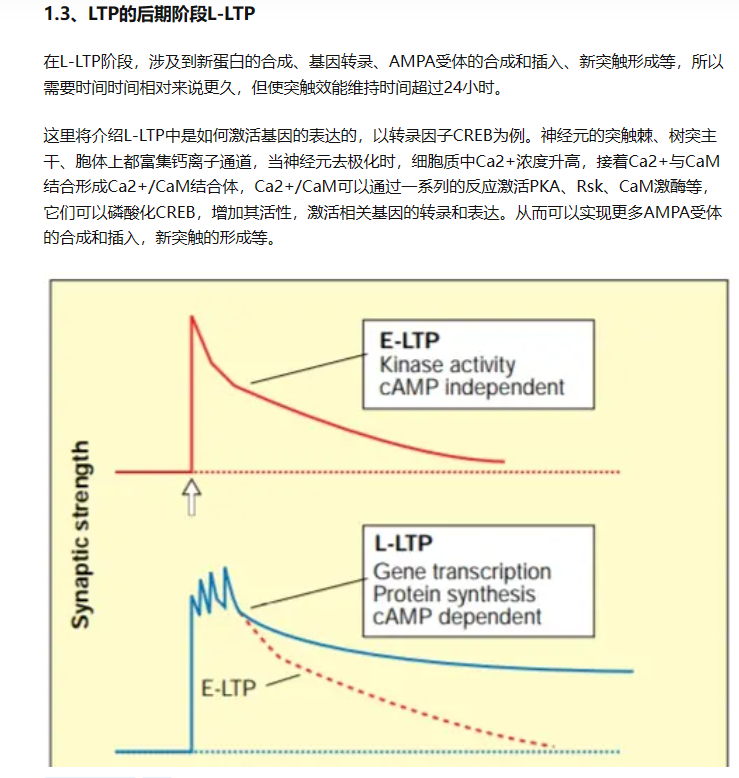
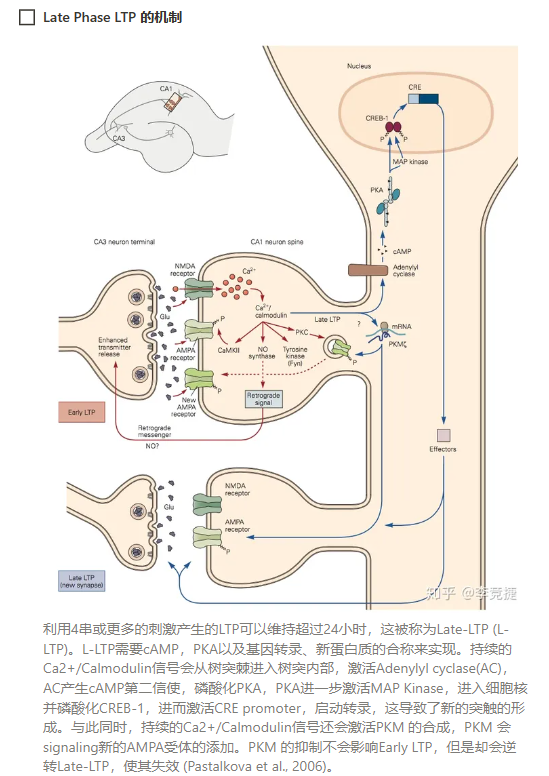

- ——大部分的LTP产生在突触后神经元(原理:AMPA受体的上膜、NMDA也在LTP中起关键作用,无NMDA则无法产生LTP——NMDA受体影响了AMPA的上膜——钙离子低于平衡电位内流导致的结果)
- Spike timing:Pre和post动作电位时间很近,且符合先来后到的原则,则是LTP
- LTP包含蛋白质的合成
- LTP与学习记忆相关:NR2B的突变导致小鼠变聪明(LTP的延长)、条件化记忆的实验(小鼠的电刺激)、小鼠的空间记忆(NMDAS受体的突变)