神经科学简史

- 皮肤的感觉信号和肌肉运动的信号是否通过同一根神经电缆?如何验证?

- Broca区(语言的发音)、韦尼克区(理解语言)
- 神经系统形态的研究方法:
- 尼氏染色:

- 高尔基染色:
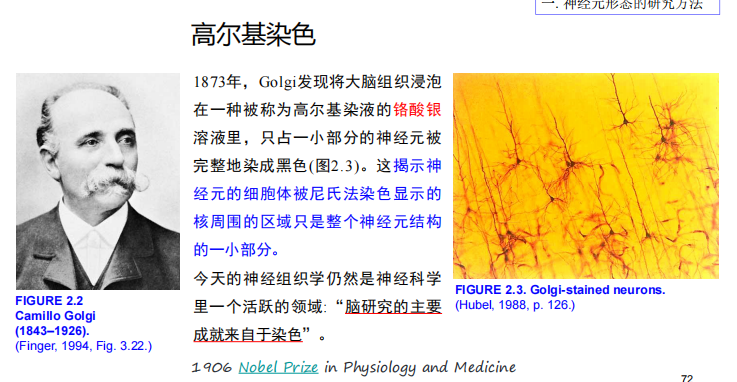 注:Cajal用Golgi 染色法染出了许多脑区的环路
注:Cajal用Golgi 染色法染出了许多脑区的环路
- 尼氏染色:
- 20世纪神经科学的进展:
- 突触的发现
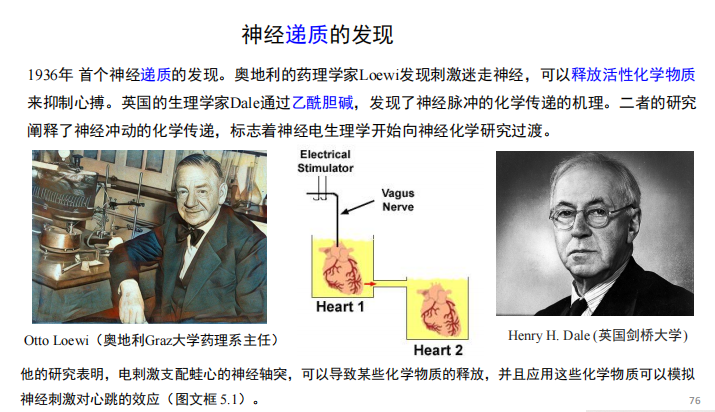
- 神经递质的发现:
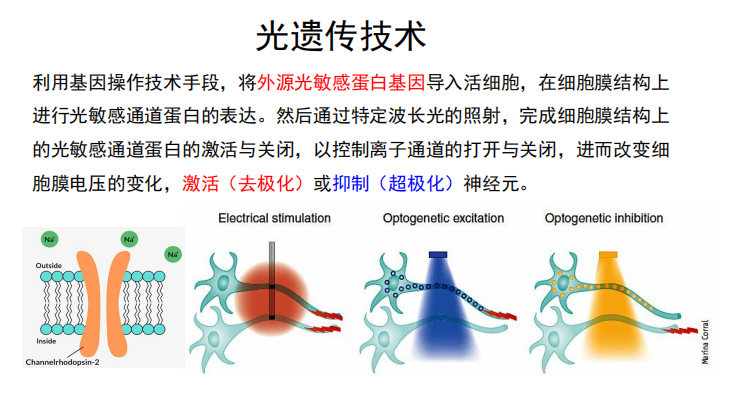
- 单神经纤维的高度分化能力以及白质切除术:

- 裂脑人:左右脑的协调分工
- 神经生物学的内容和研究层次:
- 还原法(分子、细胞、系统、行为、认知):
- 分子水平:神经系统的基因、蛋白质、及其他分子的结构与功能
- 细胞神经科学:
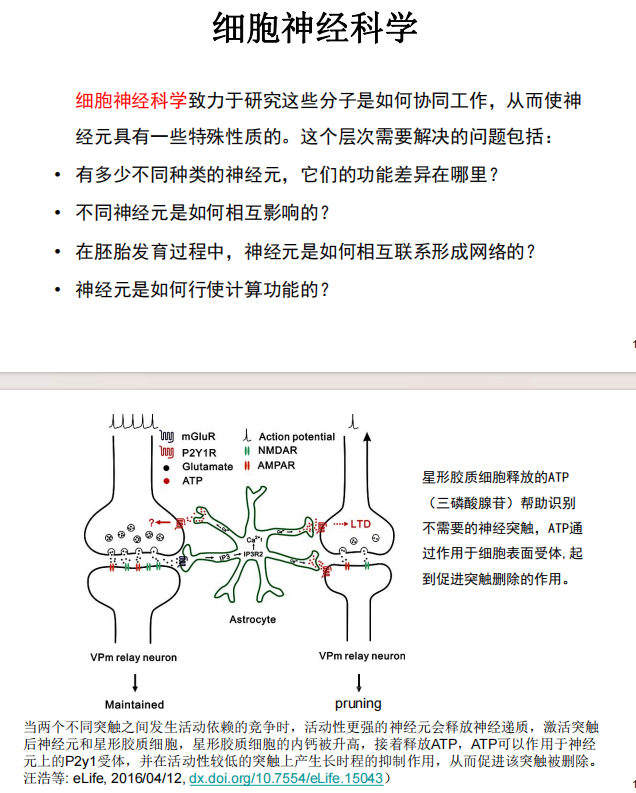
- 系统神经科学:环路

- 行为神经科学:

- 认知神经科学:

- 分子水平:神经系统的基因、蛋白质、及其他分子的结构与功能
- 谈一谈脑与认知科学的发展趋势:

- 还原法(分子、细胞、系统、行为、认知):
脑的基本结构及其二态性
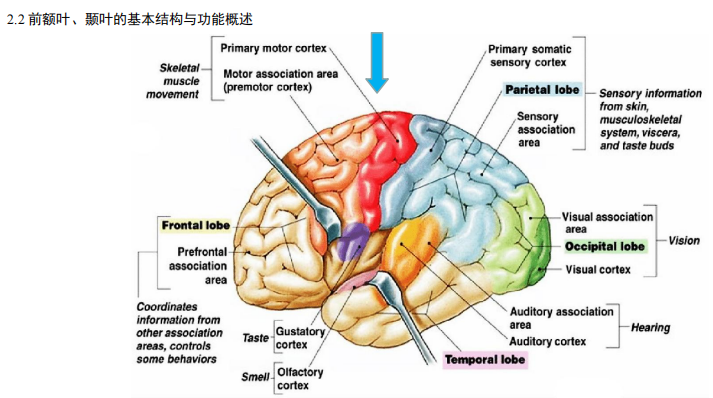
- 前额叶、颞叶的基本结构与功能概述:
 注:人脑厚而复杂的外层部分归功于NOTCH2NL基因家族。
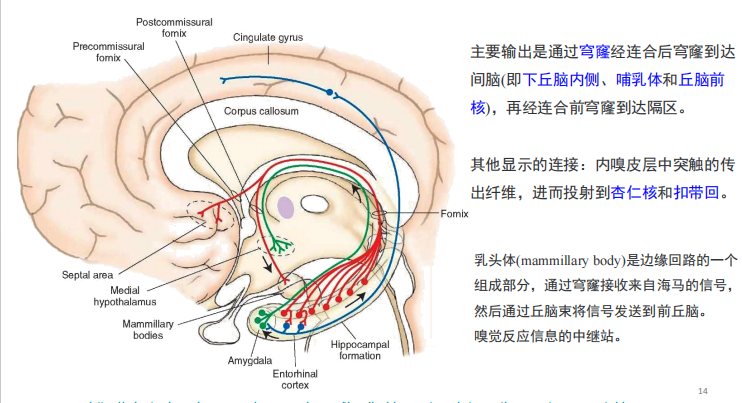
注:人脑厚而复杂的外层部分归功于NOTCH2NL基因家族。 - 边缘系统:边缘系统能将高级的心理功能和原始情感结合在一起。 它不仅关系到我们的情感生活,而且关系到高级的心理功能,例如学习和记忆的形成。 边缘系统是某些身体事物(例如进食)对我们来说如此令人愉悦的原因,也是某些医疗状况(例如高血压)是由精神压力引起的原因 重点: · 位于丘脑两侧大脑下方的边缘系统不仅负责我们的情感生活,而且还负责许多高级心理功能,例如学习和记忆的形成。 · 边缘系统内的主要结构包括杏仁核,海马,丘脑,下丘脑,基底神经节和扣带回。
 注:与陈述性记忆相关的脑区如下
注:与陈述性记忆相关的脑区如下
- 海马:
- 杏仁核:

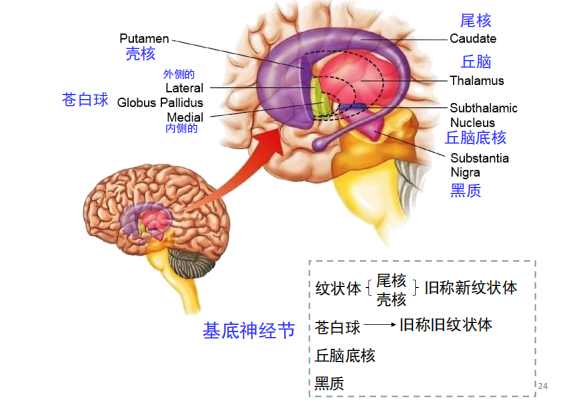
- 基底神经节:运动系统的刹车系统,与帕金森有关
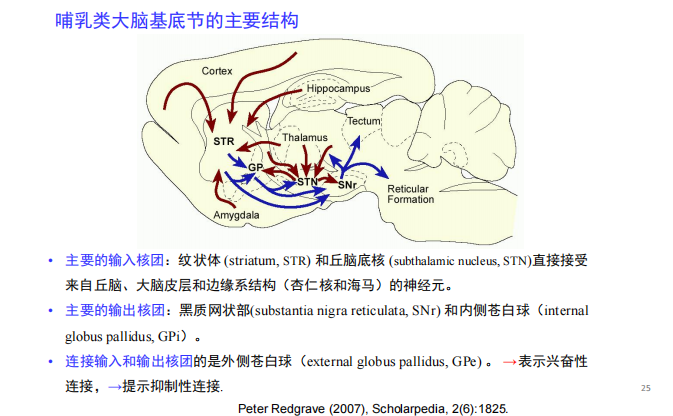
- 海马:
- 脑的二态性:
- 生殖器官及神经调节:
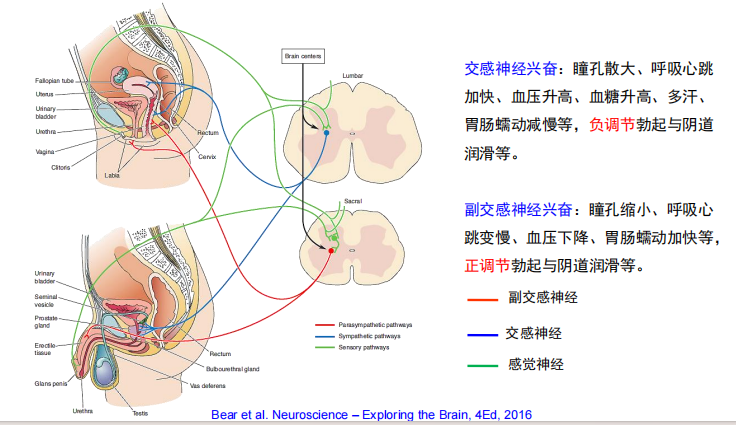 注:完成一个性反应周期需要神经系统的交感神经参加。当感觉轴突(尤其是阴茎和阴蒂的感觉轴突)活性升高时,它们和从脑下行的活动,共同兴奋脊髓胸腰段的交感神经。交感神经轴突,触发了射精过程
注:完成一个性反应周期需要神经系统的交感神经参加。当感觉轴突(尤其是阴茎和阴蒂的感觉轴突)活性升高时,它们和从脑下行的活动,共同兴奋脊髓胸腰段的交感神经。交感神经轴突,触发了射精过程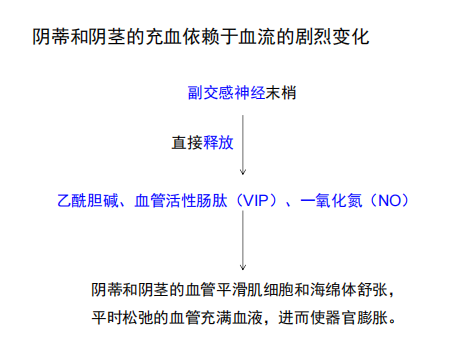 注:性高潮时,前额叶被抑制,而边缘系统表现出强烈的兴奋。
注:性高潮时,前额叶被抑制,而边缘系统表现出强烈的兴奋。 - 二态性的体现:

- 哺乳动物的交配策略:

- 雌性本能或自主地精心选择配偶
- 雄性:留下or离开

- 生殖行为的神经化学(关于抗利尿激素与催产素):


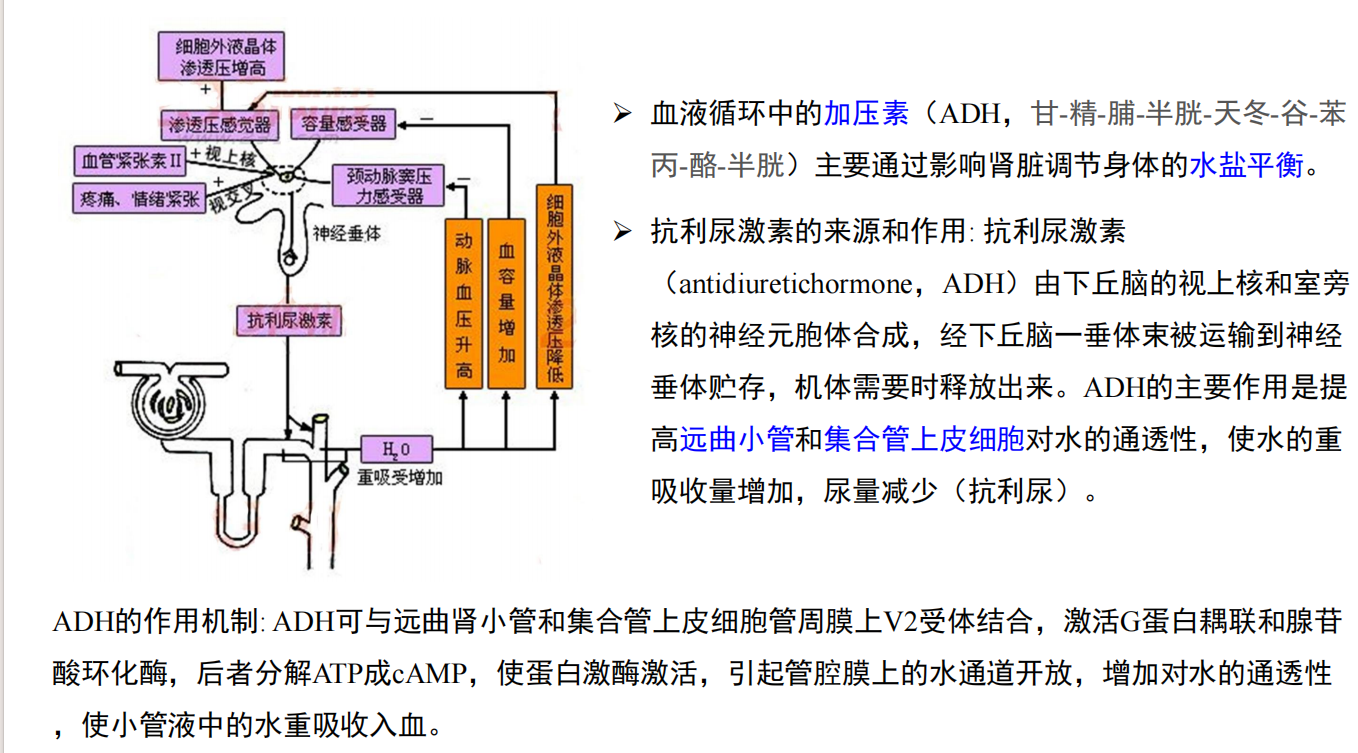
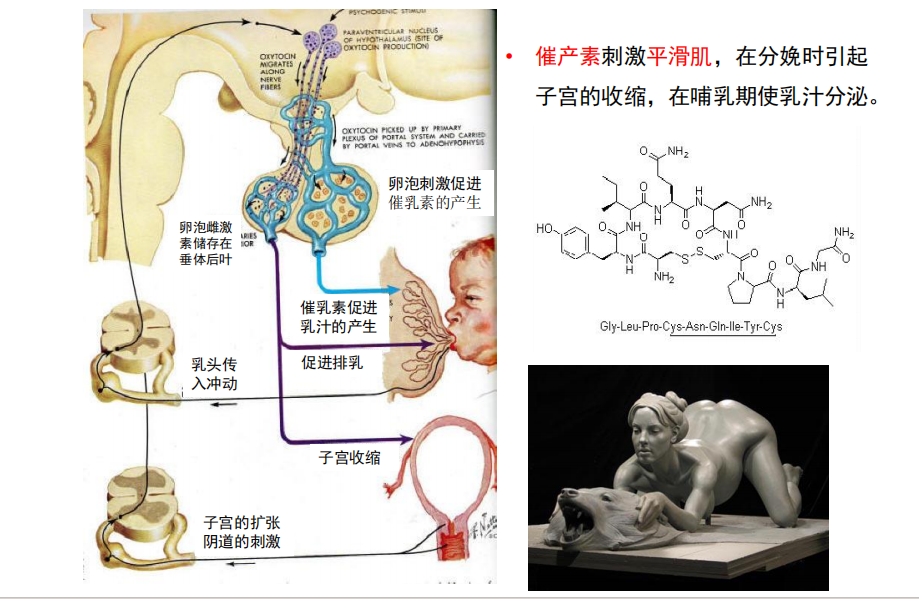 注:加压素和催产素的受体的分布在草原田鼠和高山田鼠是完全不同的
注:加压素和催产素的受体的分布在草原田鼠和高山田鼠是完全不同的

- 生殖器官及神经调节:
- 中枢神经系统性别二态性:
- 哺乳动物的视前区:
 注:如大鼠雄性脑中被贴切地称为性二态核(sexually dimorphic nucleus, SDN)的核团比雌性大5-8倍;两个性相关神经元MCMs,能帮助蠕虫寻找伴侣交配和寻找食物之间学会判断优先度。当这些神经元功能丧失,雄性蠕虫不再具有这种理解力。
注:如大鼠雄性脑中被贴切地称为性二态核(sexually dimorphic nucleus, SDN)的核团比雌性大5-8倍;两个性相关神经元MCMs,能帮助蠕虫寻找伴侣交配和寻找食物之间学会判断优先度。当这些神经元功能丧失,雄性蠕虫不再具有这种理解力。 - 人:视前区
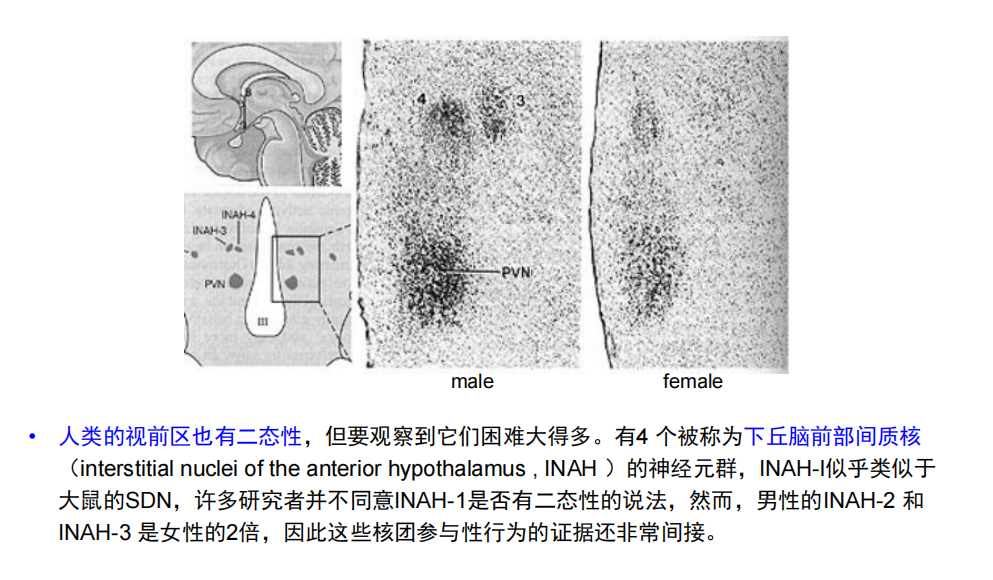 注:胼胝体不一定是;单凭核磁不可分辨性别
注:胼胝体不一定是;单凭核磁不可分辨性别
- 哺乳动物的视前区:
- 认知性别的二态性
- 性取向:5-HT






激素、神经递质与动机
- 有关动机的脑区:
 边缘系统起关键作用:
边缘系统起关键作用:
- 上行网状系统:
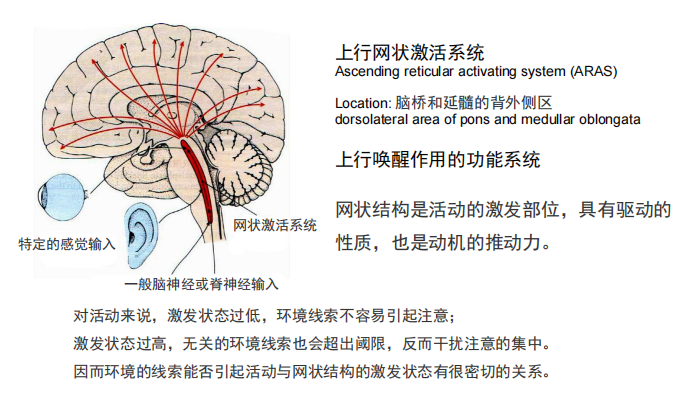
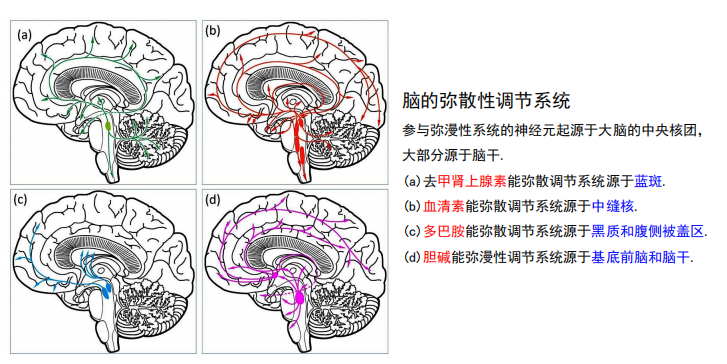
- 脑的弥散性调节系统:
- 杏仁核:

- 案例分析见思维导图:
- 男女差异:
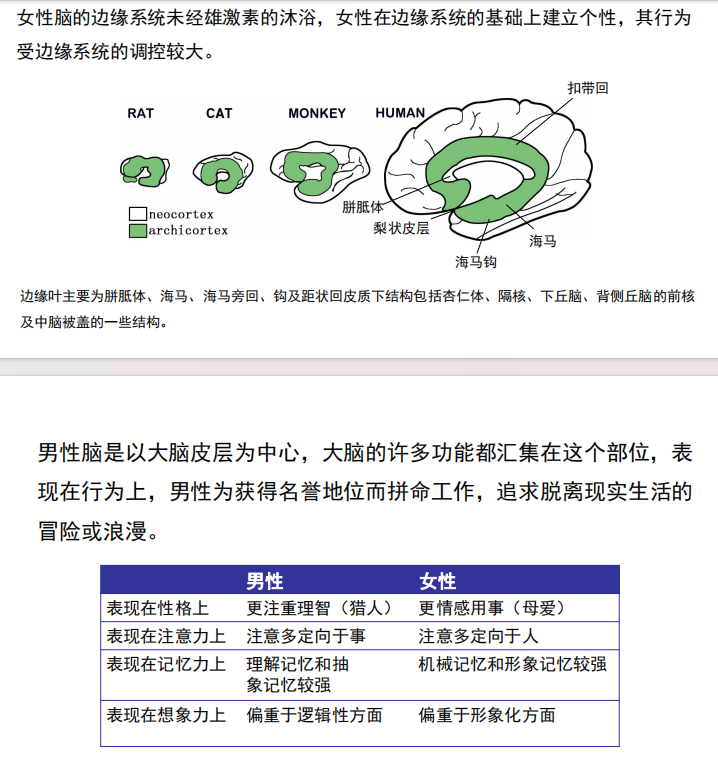
- 男女差异:
- 青春期:边缘系统与前额叶皮层之间发育不匹配,是导致青少年情绪化行为的重要原因之一
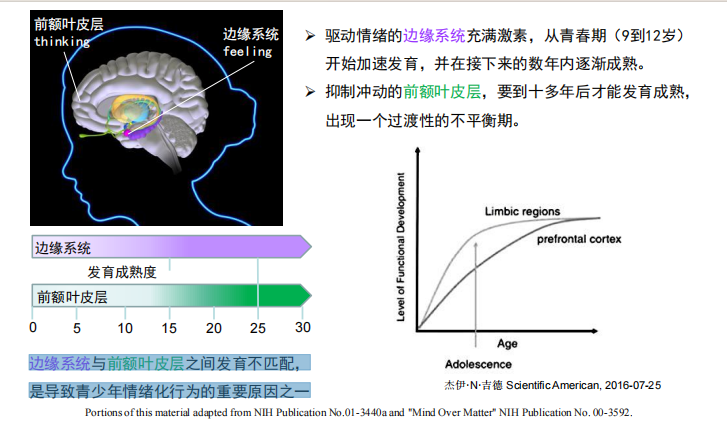
- 上行网状系统:
- 激素及受体:
- 边缘系统与动机:
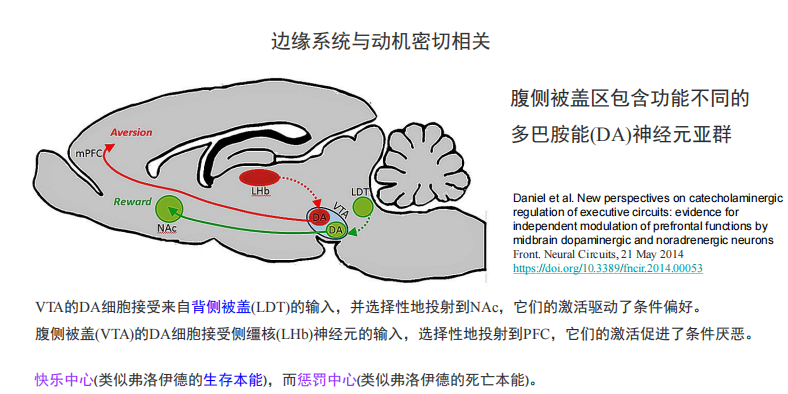
- 雄激素/雌激素与行为,见ppt第三讲108-126


视觉
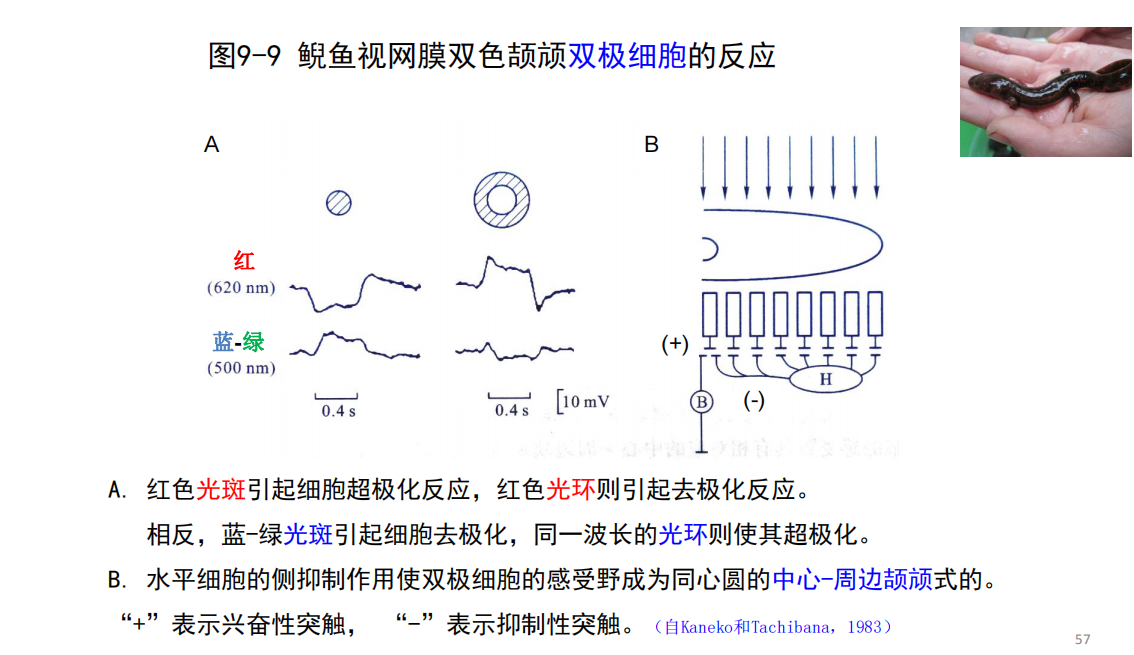
- 水平细胞感受野大?——引出了光感受器同心圆感受野的机制
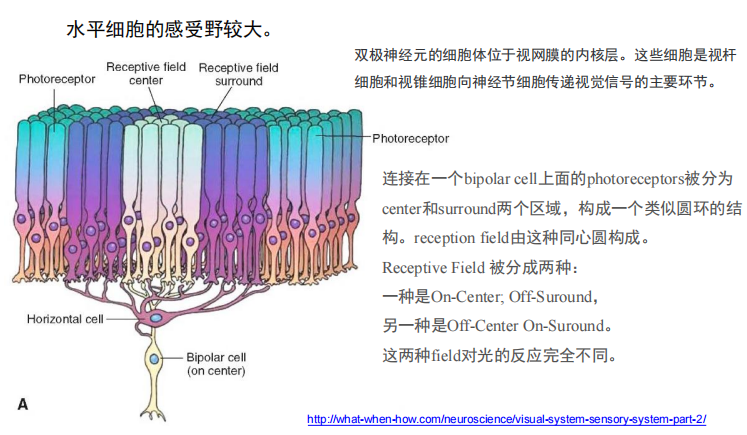 双极细胞的“中心-周边颉颃式”机制
双极细胞的“中心-周边颉颃式”机制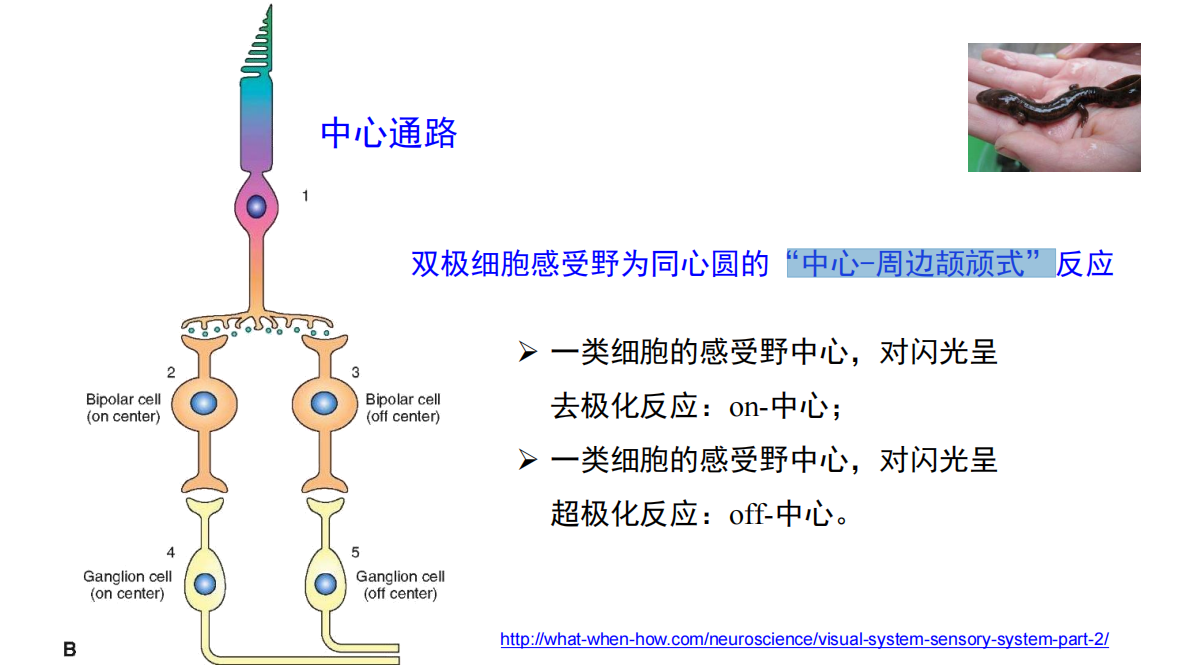
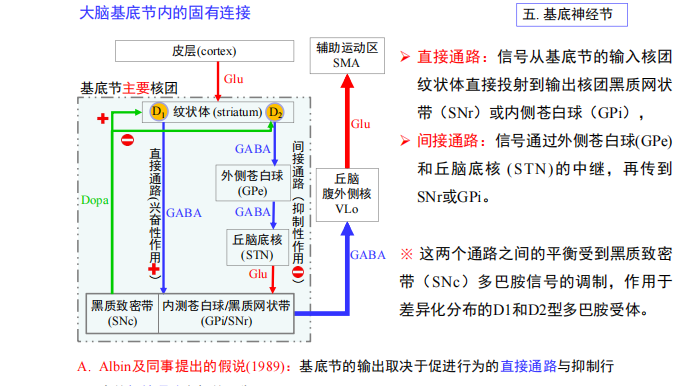
- 直接通路:
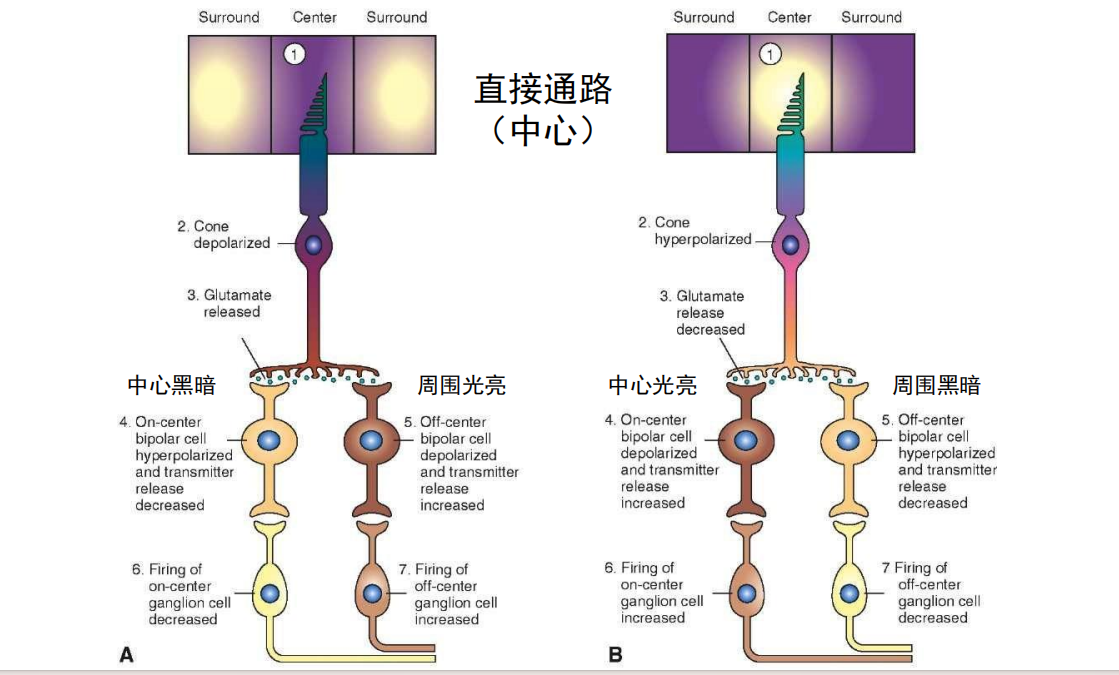
- 间接通路:
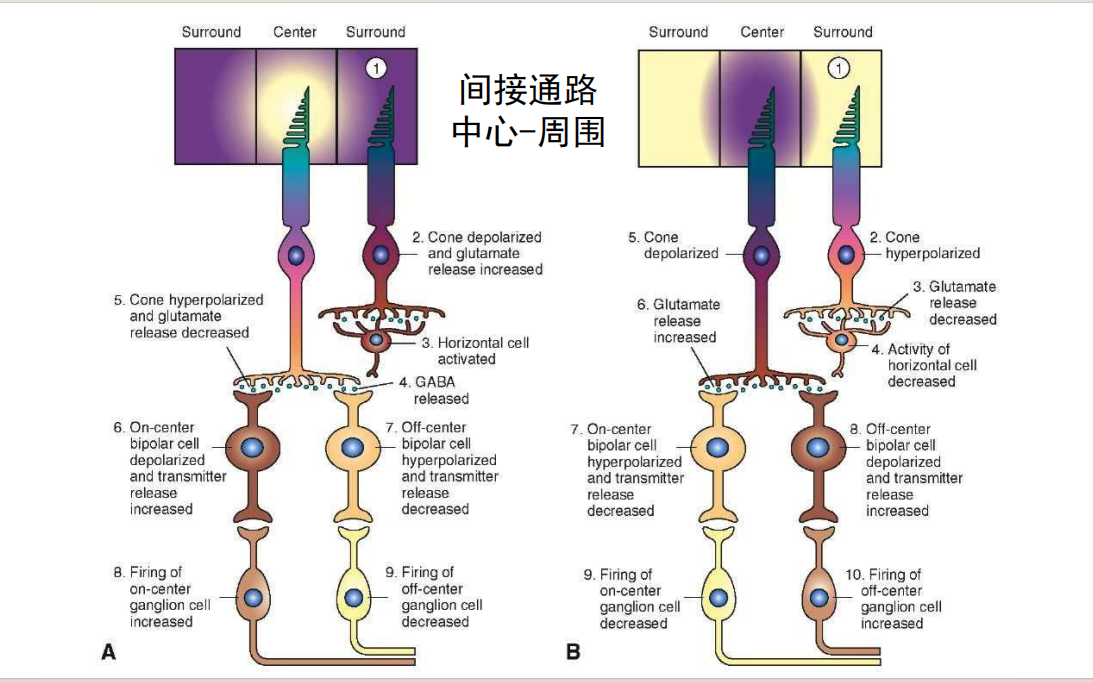
- 对颜色的on/off:
- 总结:
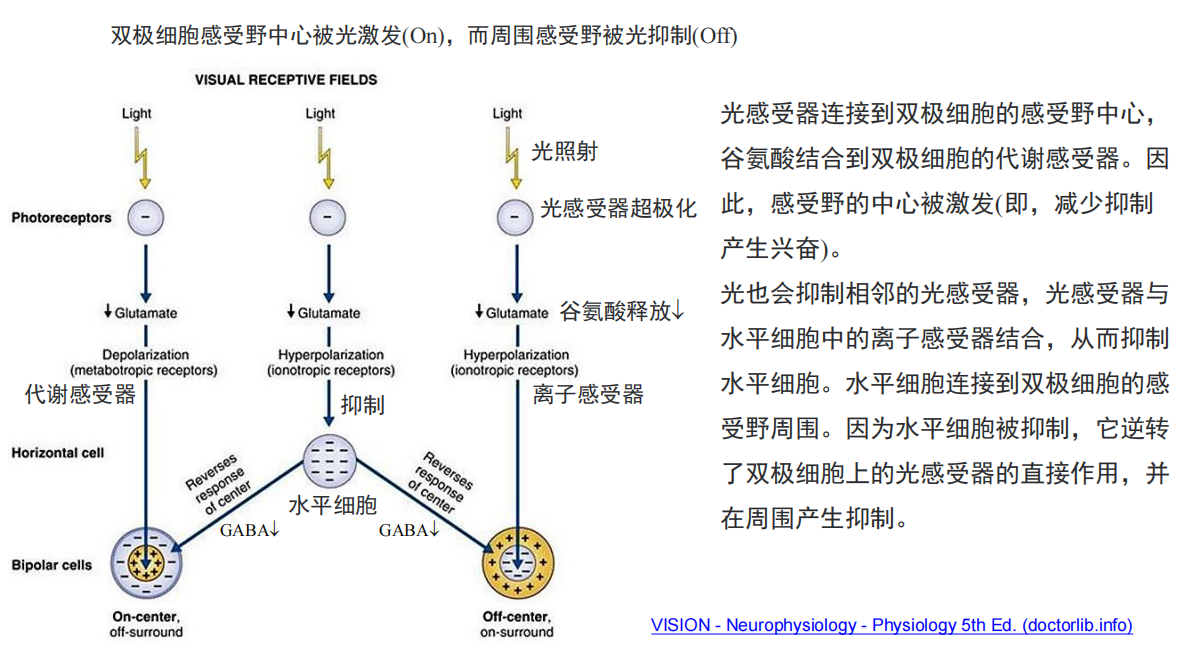
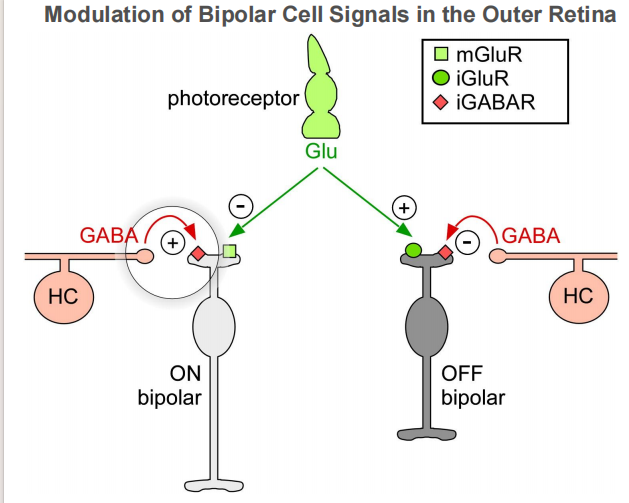
 注:当GABA由GABA能神经元释放后,与邻近神经元突触上的相应受体结合,从而最终使该神经元无法产生动作电位,那么该神经元就无法释放其突触内的神经递质,这样就不能影响与之相接触的其它神经细胞。所以说,如果GABA作用于释放兴奋性神经递质的神经元上,那么产生的最终效果则是抑制性的。反之,如果GABA和释放抑制性神经递质的神经细胞上的受体结合,那么就使该细胞无法释放抑制性的神经递质,从而产生兴奋性的作用。
注:当GABA由GABA能神经元释放后,与邻近神经元突触上的相应受体结合,从而最终使该神经元无法产生动作电位,那么该神经元就无法释放其突触内的神经递质,这样就不能影响与之相接触的其它神经细胞。所以说,如果GABA作用于释放兴奋性神经递质的神经元上,那么产生的最终效果则是抑制性的。反之,如果GABA和释放抑制性神经递质的神经细胞上的受体结合,那么就使该细胞无法释放抑制性的神经递质,从而产生兴奋性的作用。 注:当中心和周围接受光照有差异时,ganglion cell输出的信号最强。
注:当中心和周围接受光照有差异时,ganglion cell输出的信号最强。
- 直接通路:
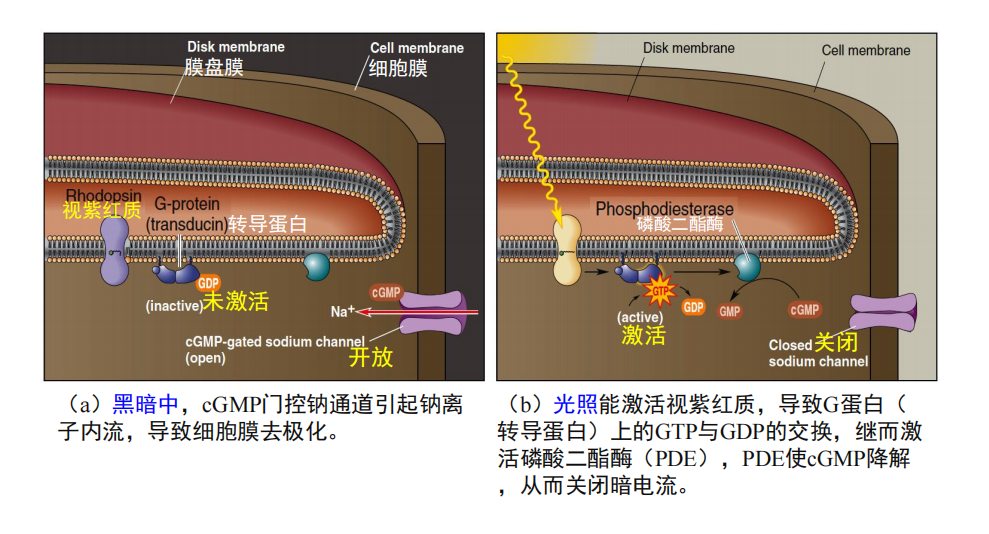
- 视觉光转换机制:

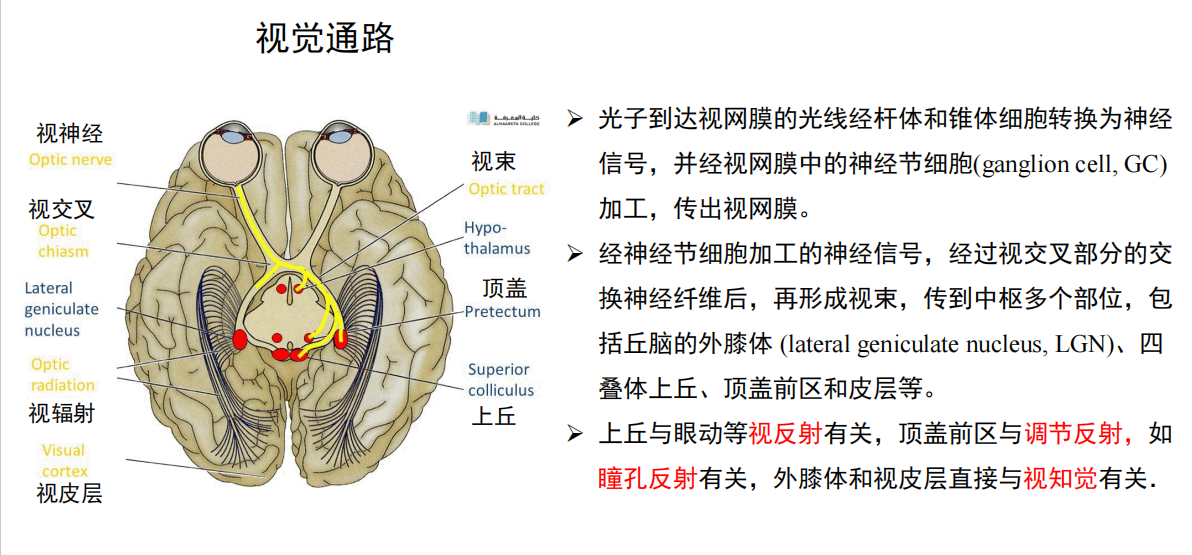
- 人类视觉系统:这说明了视觉性息也是从receptor由内到外传递的
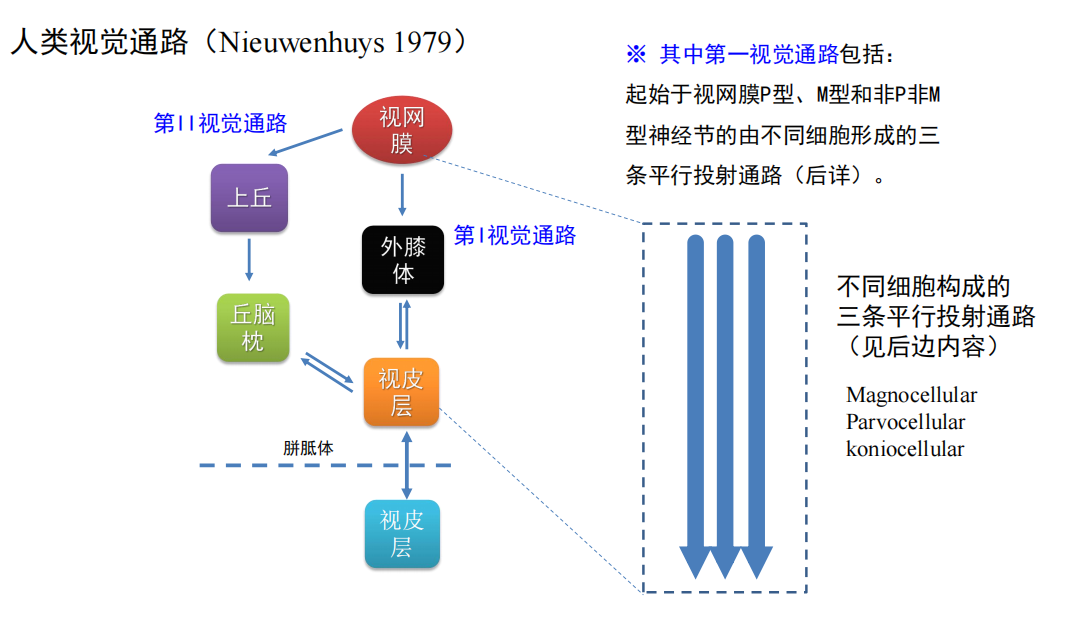
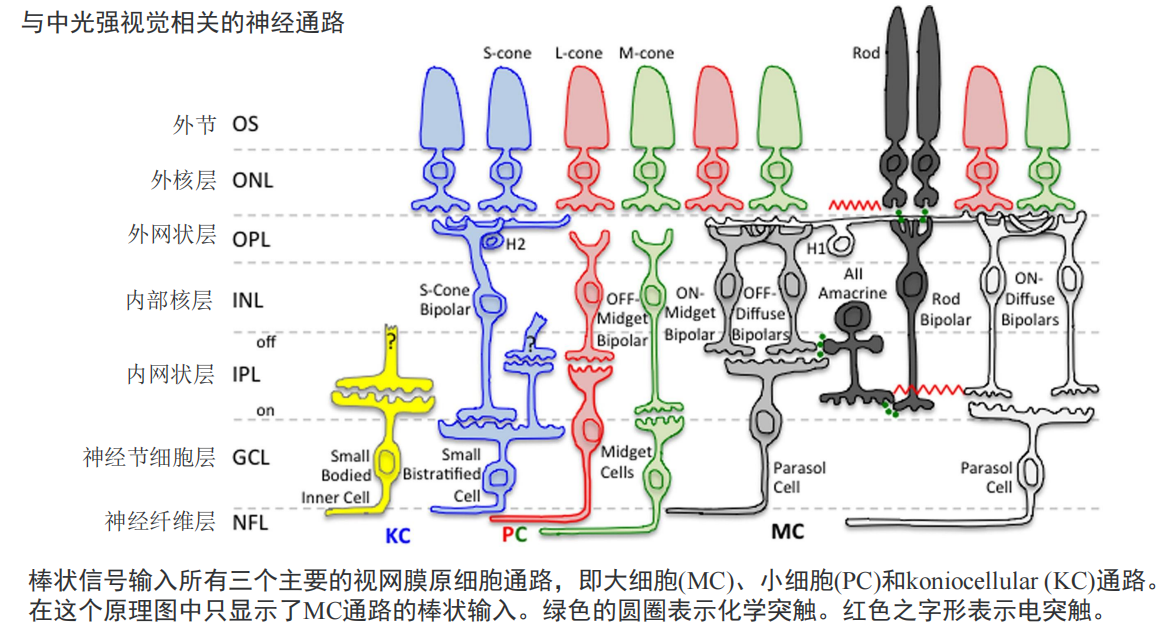 注:视觉皮层的两条通路:
注:视觉皮层的两条通路:
- What:

- Where:
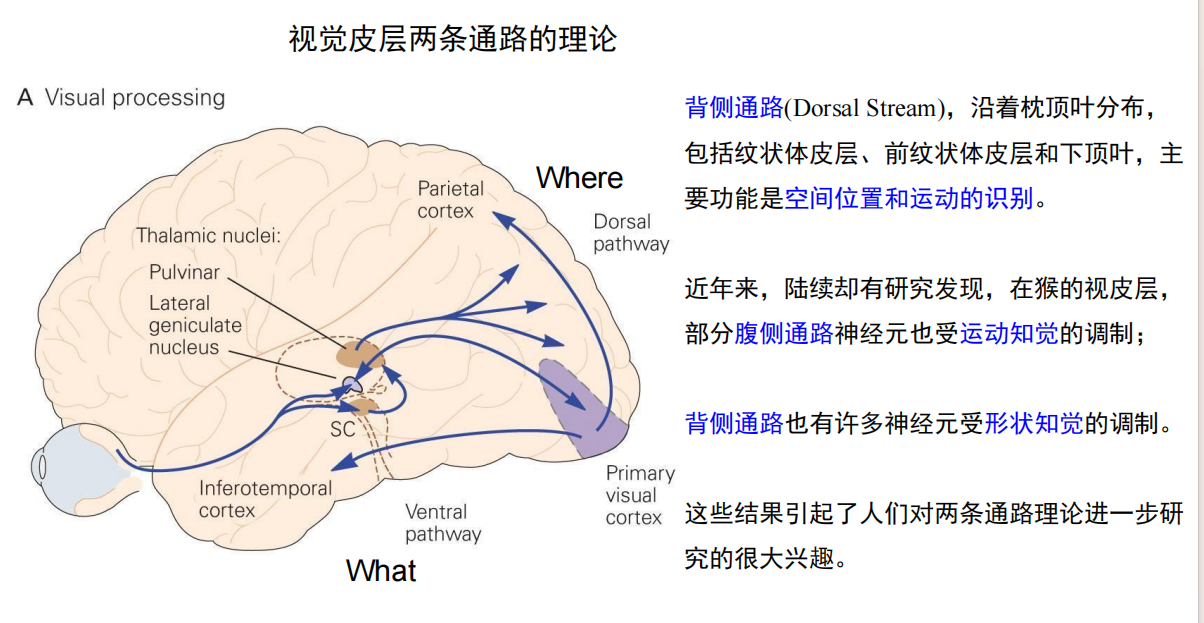
- What:
- 视觉系统的前馈模型:点线面
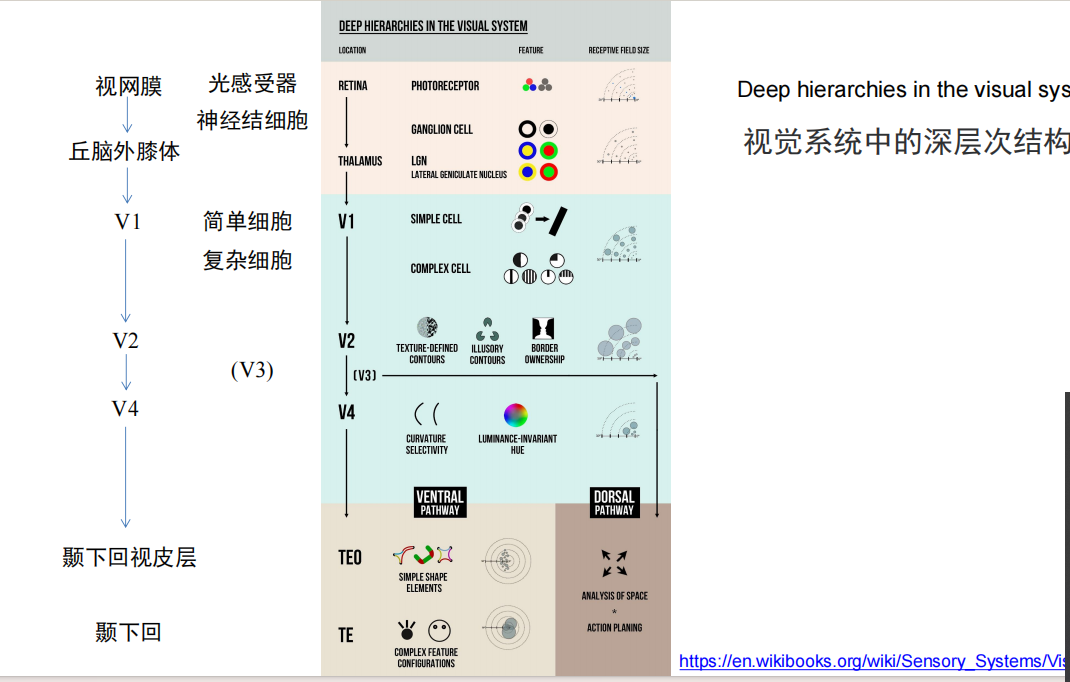
- question:
- 一次自行车事故使人不能看见左侧半视野中的任何物体,请问离视网膜通路中的哪个部位收到了损伤?——右侧视束
- 左侧外侧膝状体的主要输入来自何处?——视网膜的角度答
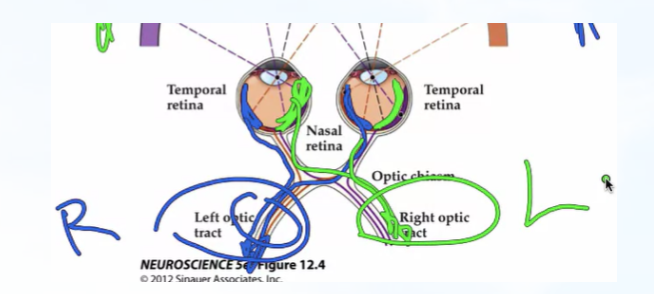
- 一侧外侧膝状体核受到损伤使人丧失了右眼右侧视野的颜色感知.请问哪个外侧膝状体核的哪个细胞层受到了损伤?
 ——右侧外膝体的235细胞层
——右侧外膝体的235细胞层 - 请列出自视网膜视锥感受器至纹状皮层斑块细胞的连接通路。

- 自视网膜M型请问哪个通路向纹状皮层提供的输入更多,是大细胞通路还是小细胞通路?这个通路主要含有对视觉世界的哪两种分析功能?其它通路呢?

- 神经节细胞至纹状体皮层IV B层神经元的连接通路中,感受野在各个突触连接处是如何转换的?

- 试述视觉系统并行处理机制的含义,并给出两个例子。

- 如果一个婴儿患有先天性内斜,并且未在10岁前进行矫正,那么它的双眼深度感知将永久性地消失。这可以用视觉系统中回来的调节加以解释。根据你掌握的关于中枢神经系统的知识,你认为这种回路的调节发生在视觉系统的哪个部分?

- 为什么说MT区比IT区更特化为对视觉运动的检测?

- 多年来,人们认为深度感知涉及每个眼睛对物体的识别,以及随后的双眼整合。那么图文框10.4所讨论的立体图是如何证明这个这个假设的?大脑的哪个部分是产生双眼整合的可能部位?

- 哪些因素影响了视锥和视杆细胞的功能差别?
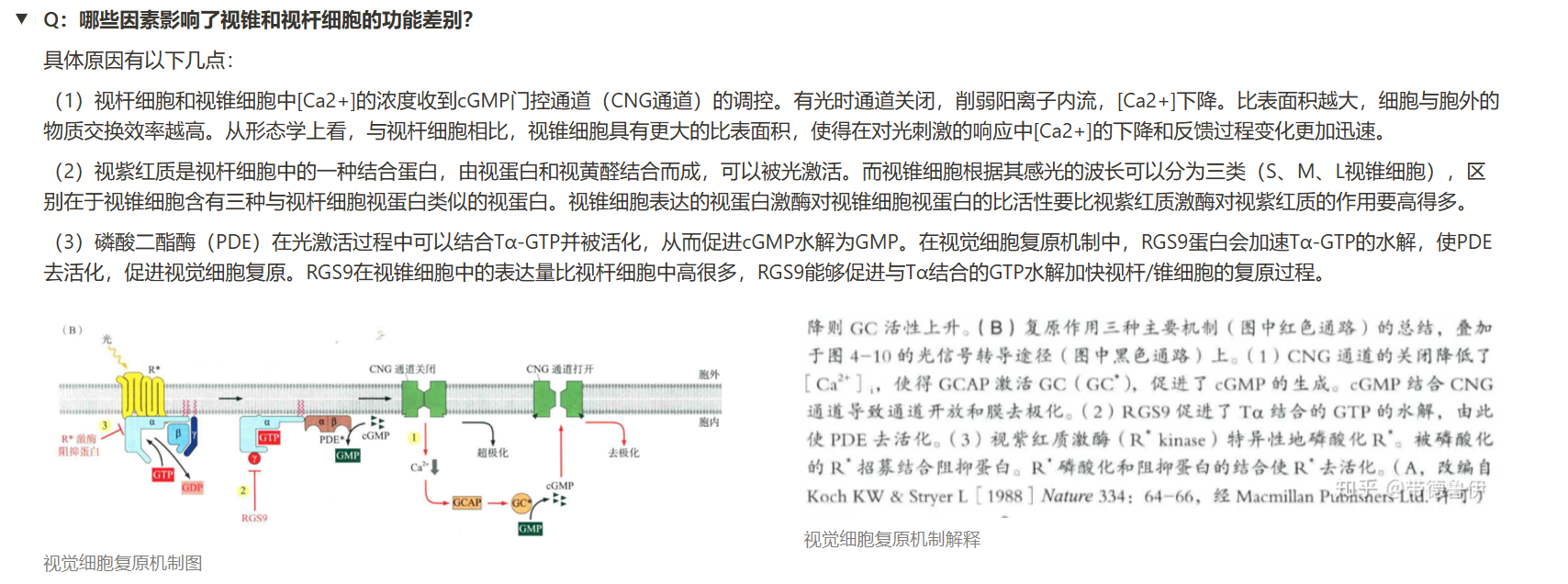
- 视锥细胞会不会产生“不应期”?

- On型、Off型RGC细胞的定义是依据什么来定义的?

- 为什么Off centered RGC神经元在外周撤光之后firing rate也增加了(如13图蓝色箭头所指)?

- 如何理解On-Off细胞的形成机制?On-Off细胞的“拮抗”是如何体现的呢?

- RGC的分类和各种RGC(视网膜神经节细胞)的分布?

- 人的RGC能够提取所有外界的信息吗?

- 那么大脑的主动控制是发生在RGC还是进入大脑以后的通路中呢?以运动的detection为例,是抑制了其他RGC还是传到大脑中再处理?——视觉高级皮层有下行调控的神经元,在外膝体处,皮层和脑干的反馈投射远大于视网膜的上行投射。但是视网膜不接受来自皮层的下行调节。
- 为什么人脑能区分背景和运动的物体?

- 交叉投射有什么意义?视觉信息为什么要投射到对侧半球进行处理?
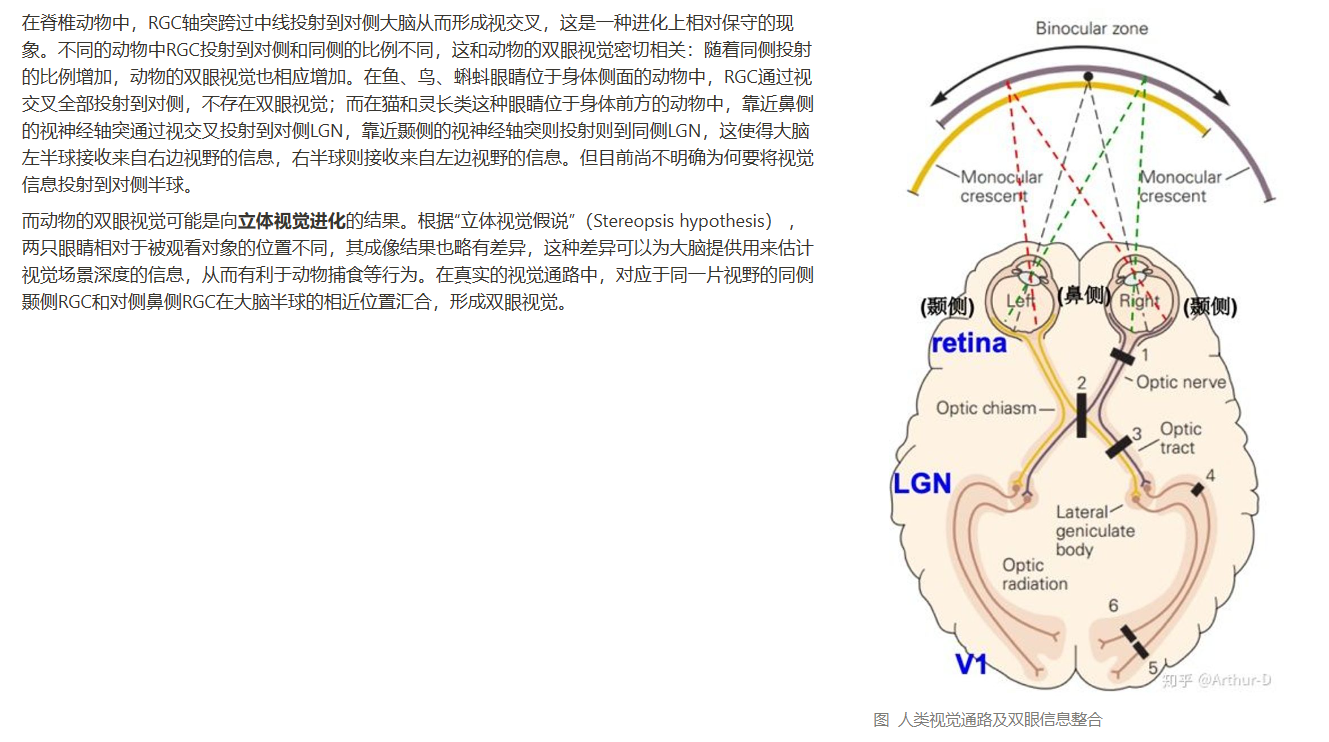
- 考虑到神经元只有一个轴突,如果LGN只有一个下游神经元,那么为了编码视觉信息的不同特征,我们就需要在相同的感受野有对应特征个数的LGN细胞。这是否比较低效?

- “背景效应”(Contextual effect)的神经机制是什么?

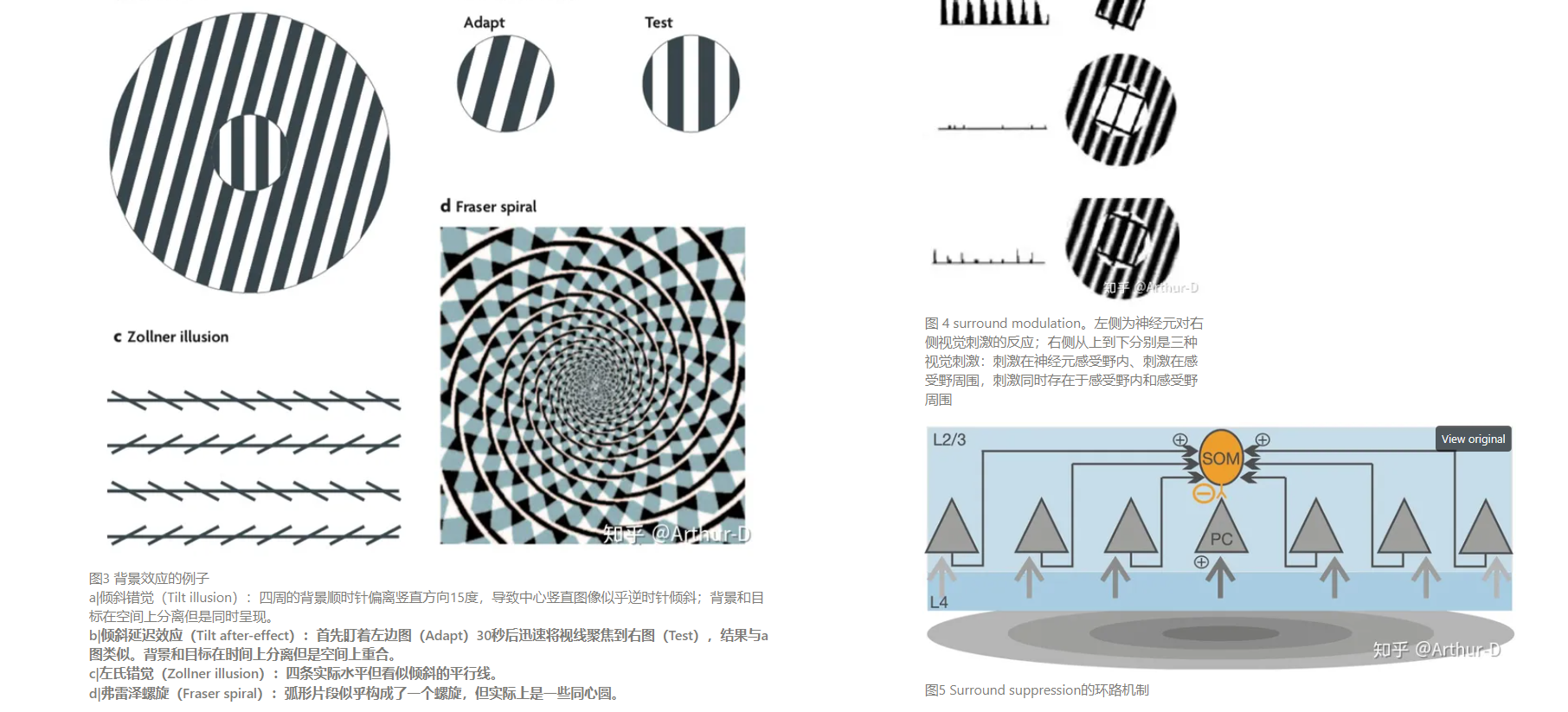
- 人们在回忆时,脑海里常会浮现出画面,这些画面的产生是兴奋了视网膜还是兴奋了视皮层?还是其他原因?

神经元与胶质细胞
见pad笔记
- 线粒体相关的神经退行性疾病:
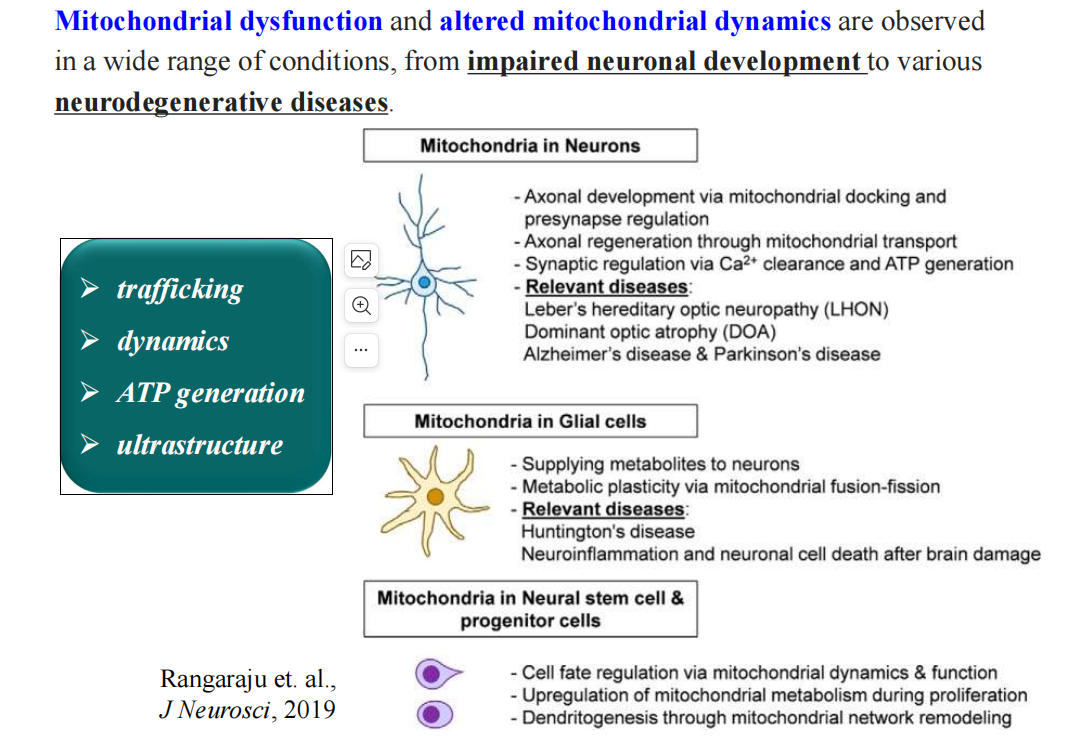
- 轴突末梢的特点:

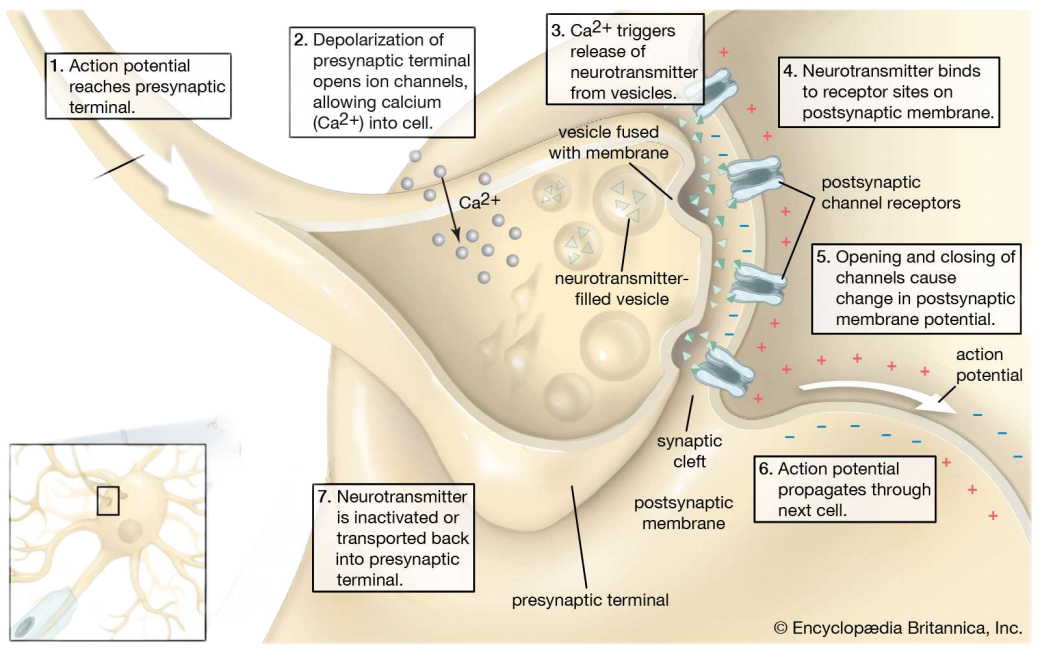
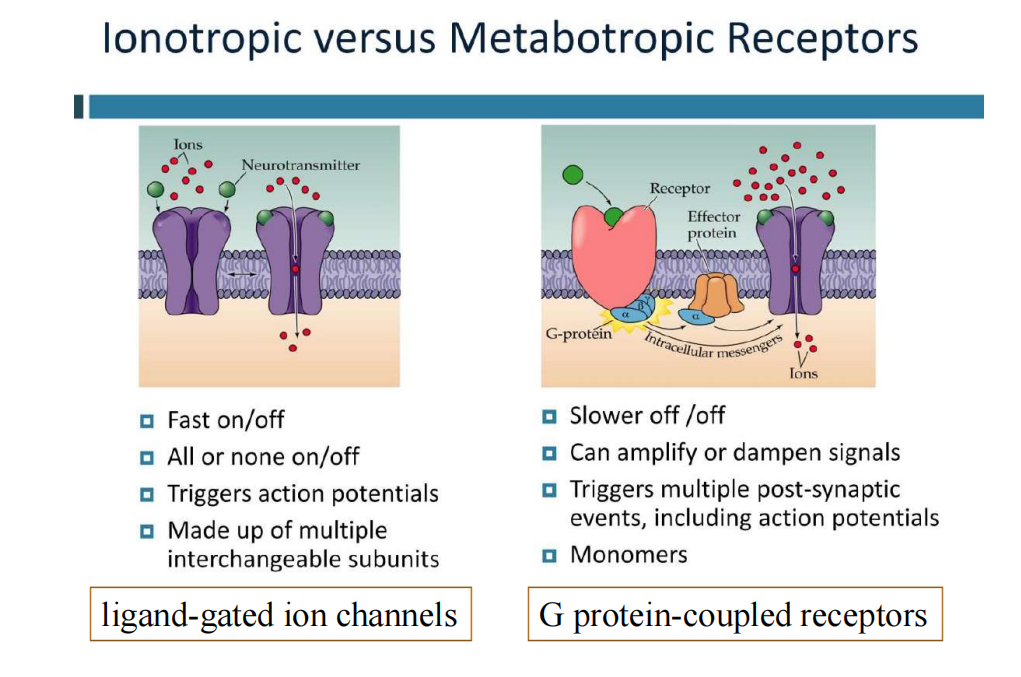
- 突触的信号传递机制以及通道:
脑和行为的化学控制——自主神经和弥散神经系统
-
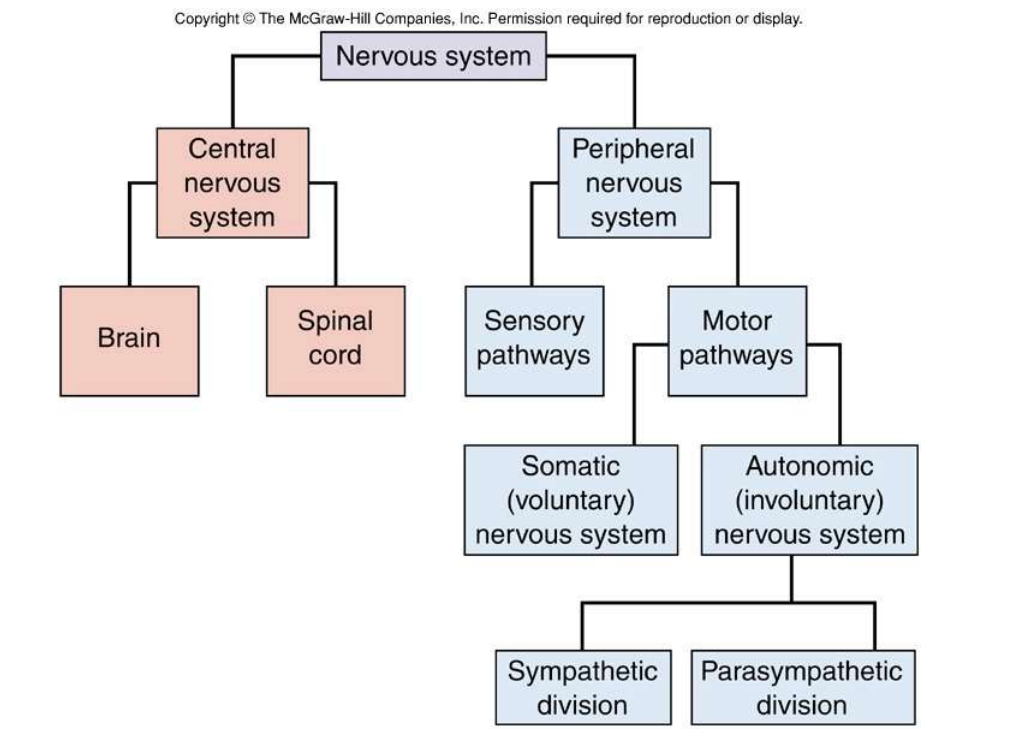
神经系统概述:

-
ANS与somatic NS对比—ppt24-28

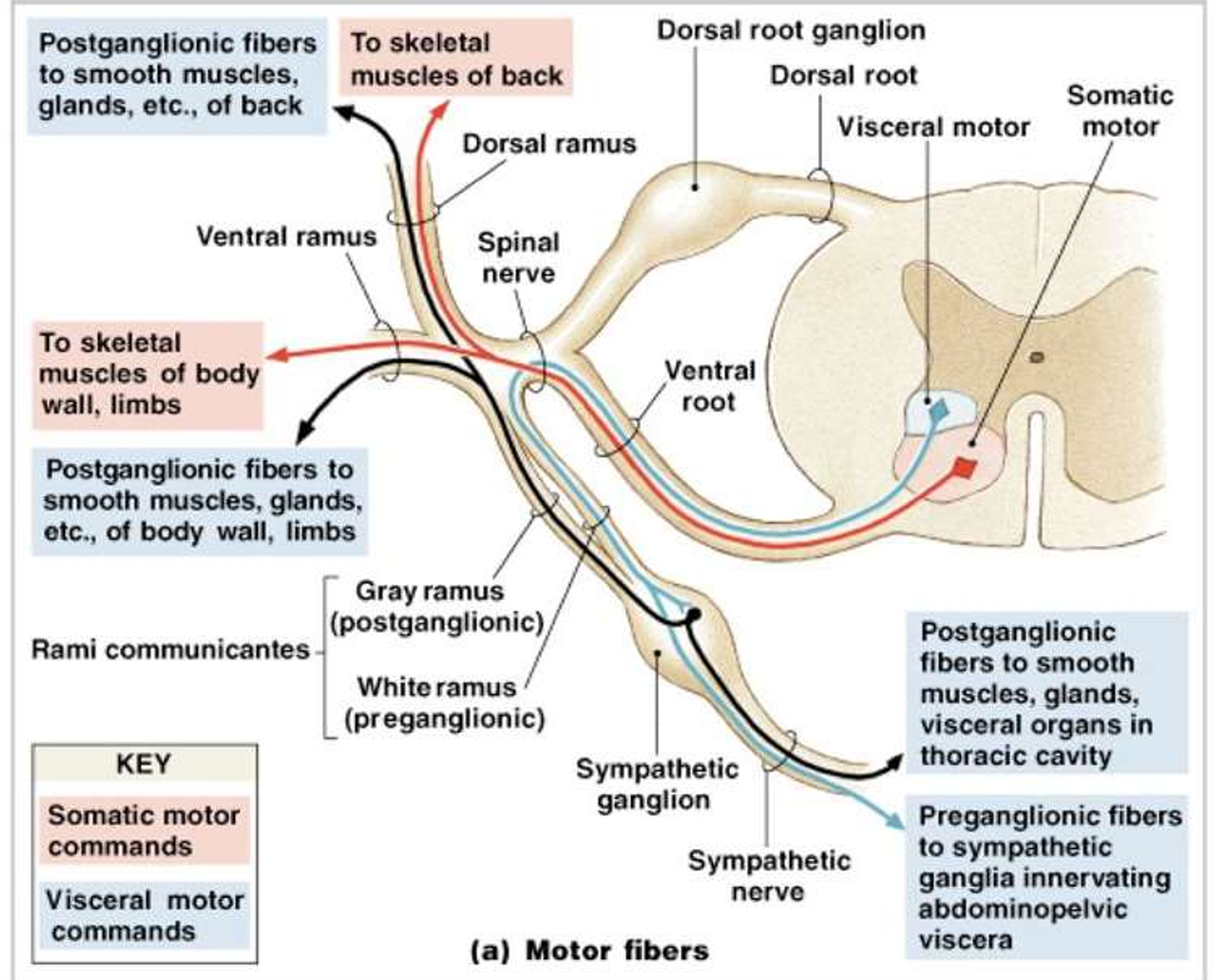 注:运动控制如上
注:运动控制如上
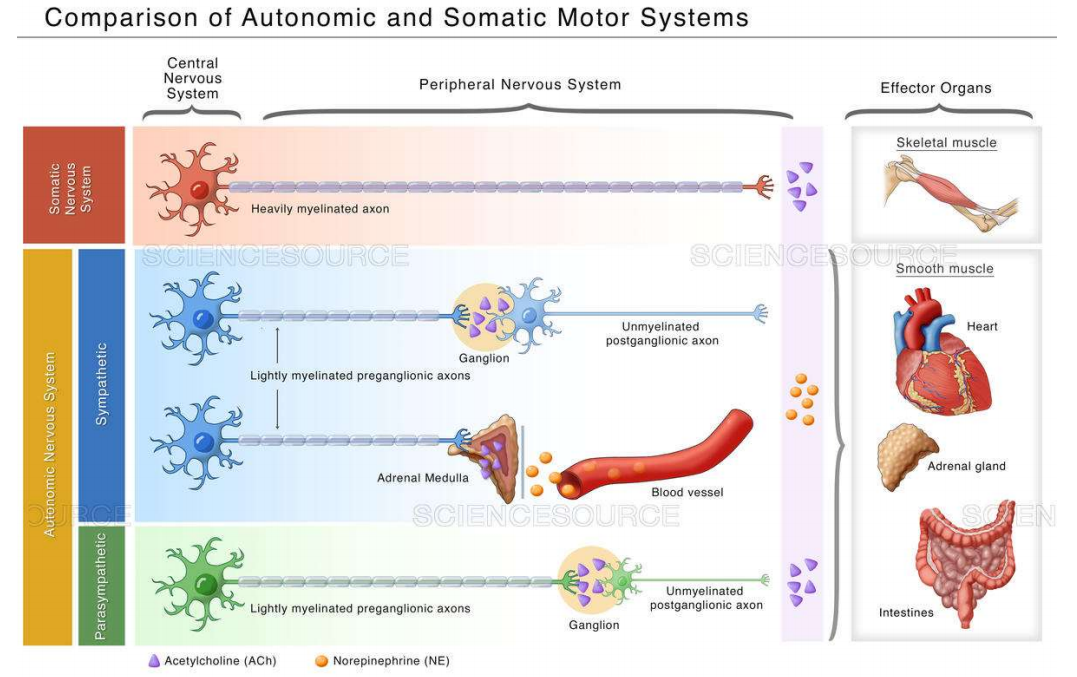
-
交感与副交感:PPT31
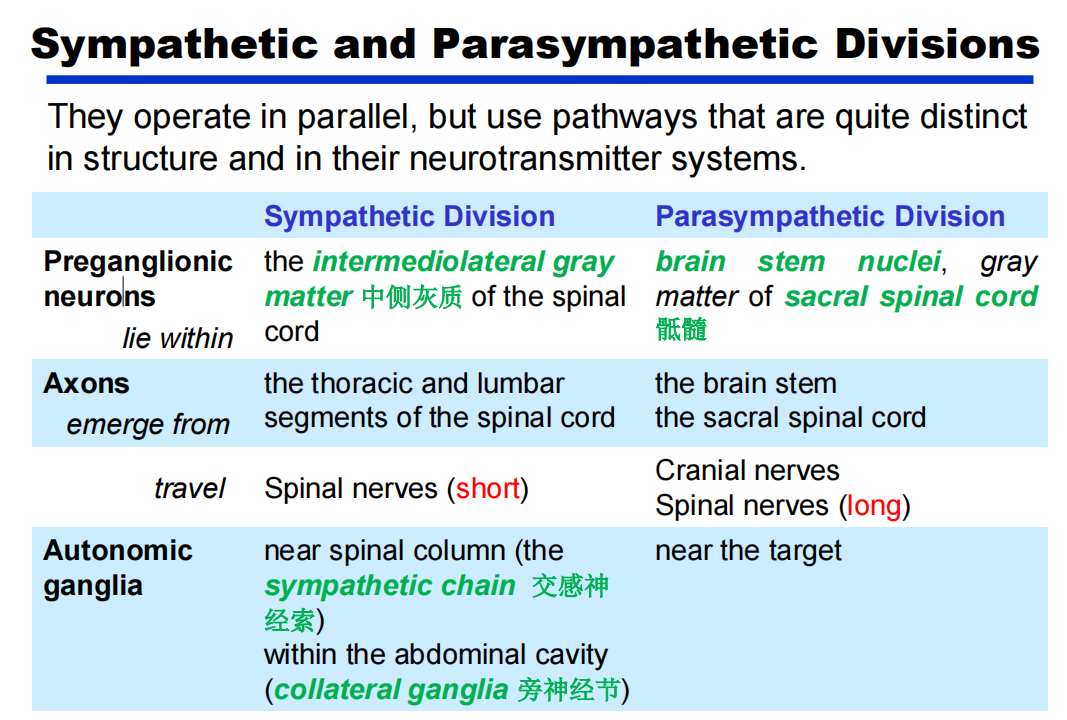
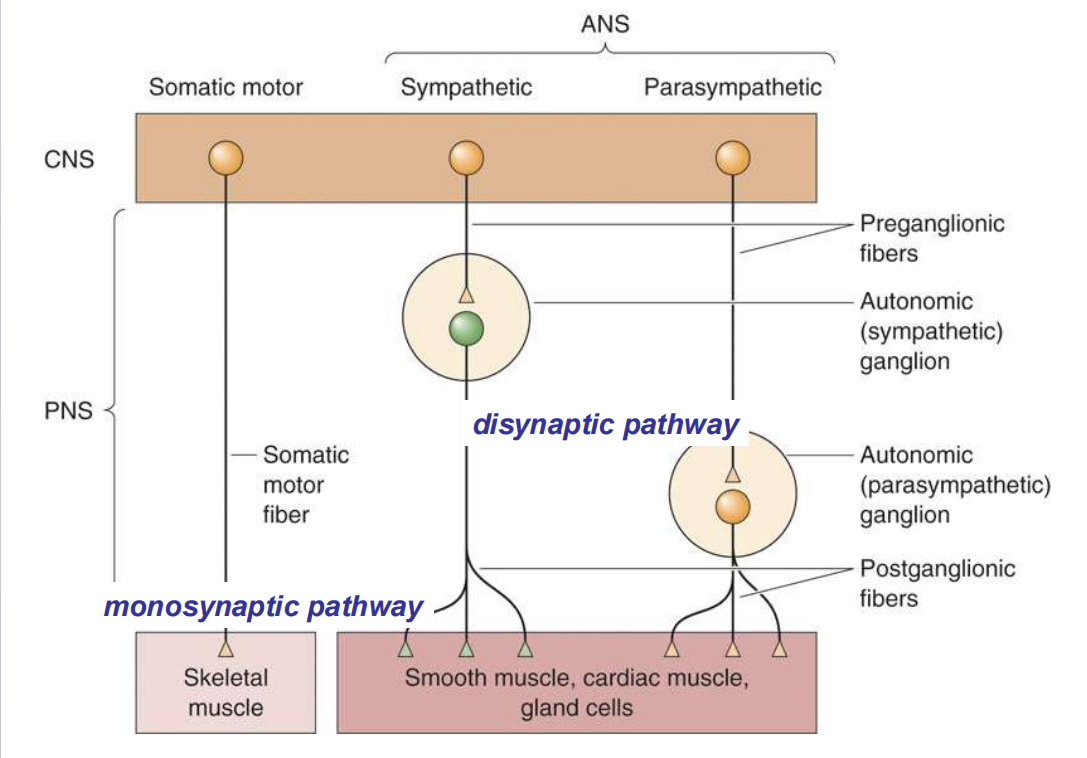
-
交感和副交感通路的信号机制:
- 节前:除了Ach还有VIP、NPY(神经信号肽类—ppt42)

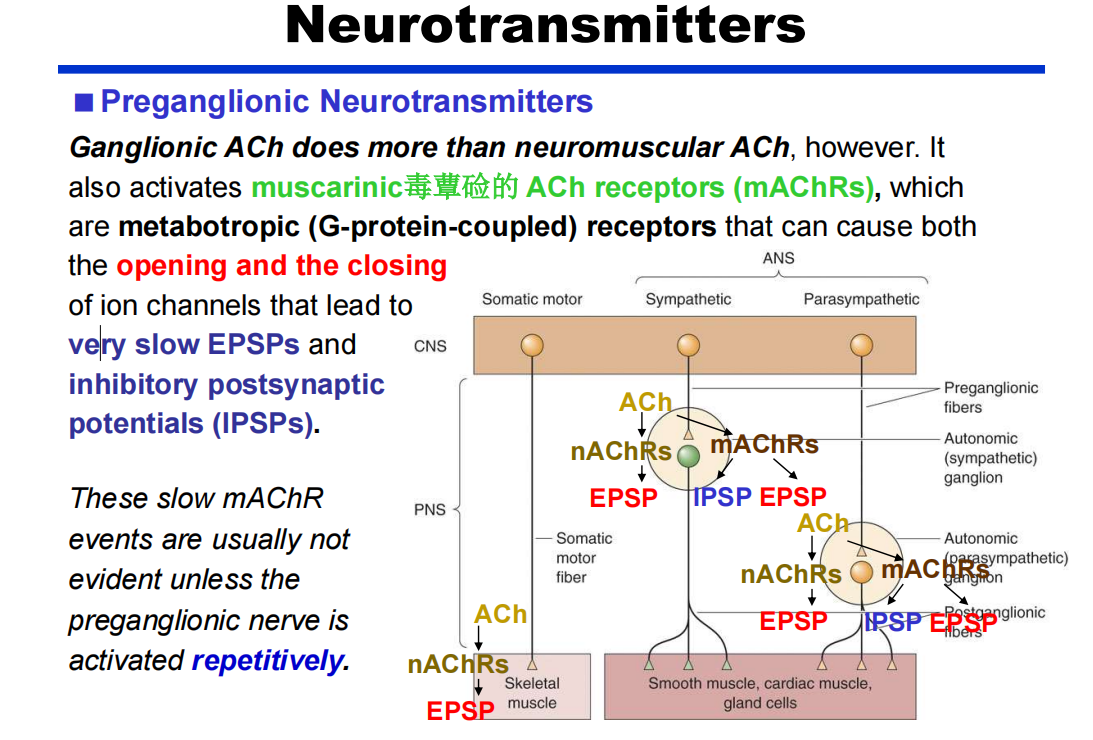
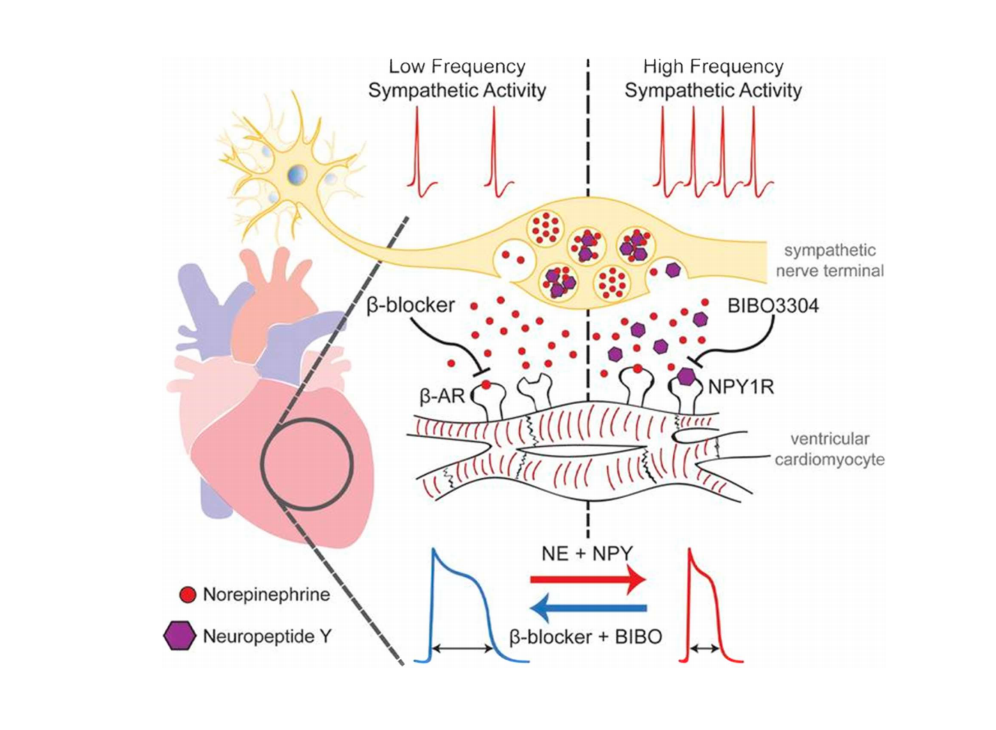 注:节前神经元放电模式的改变会影响释放物的不同
注:节前神经元放电模式的改变会影响释放物的不同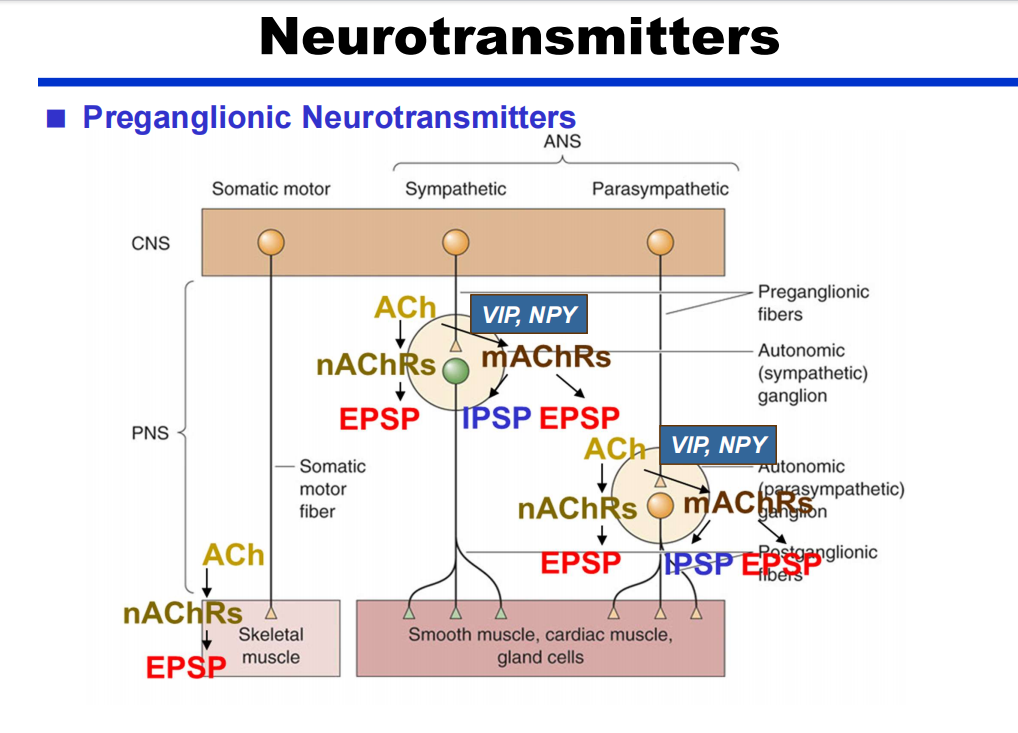
- 节后:
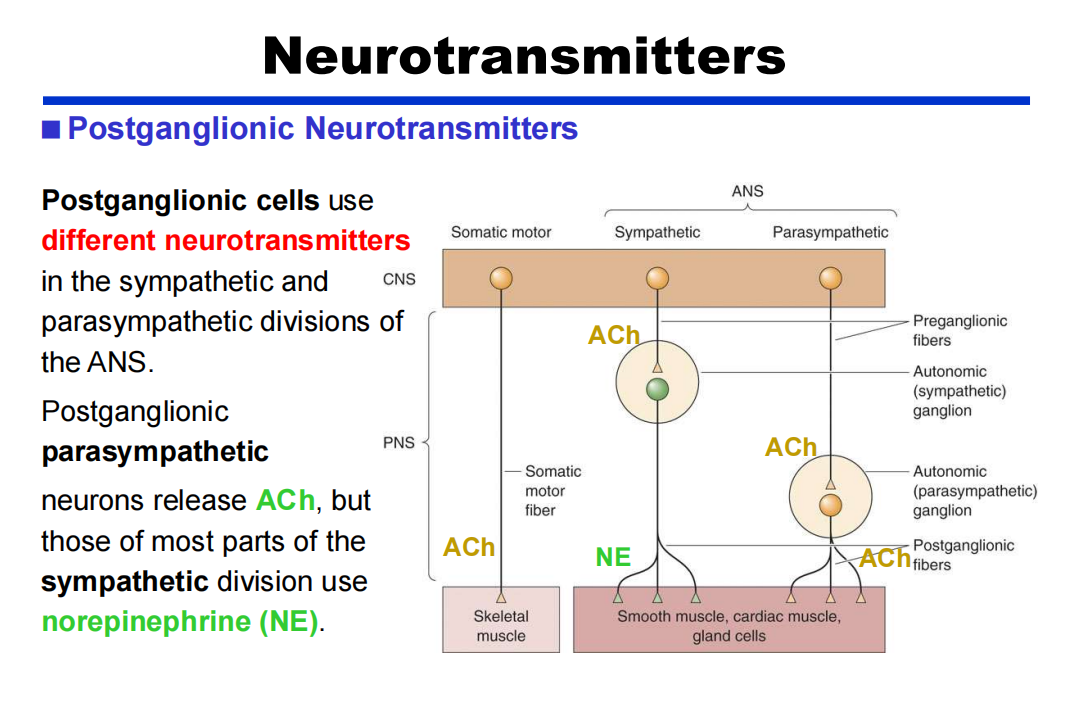
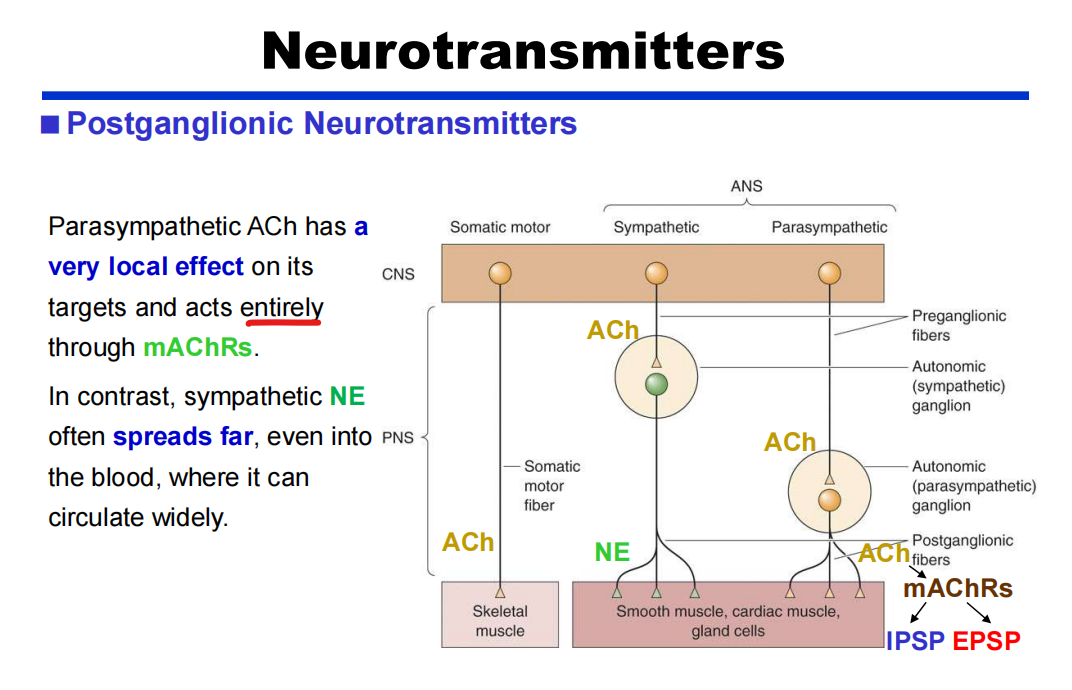
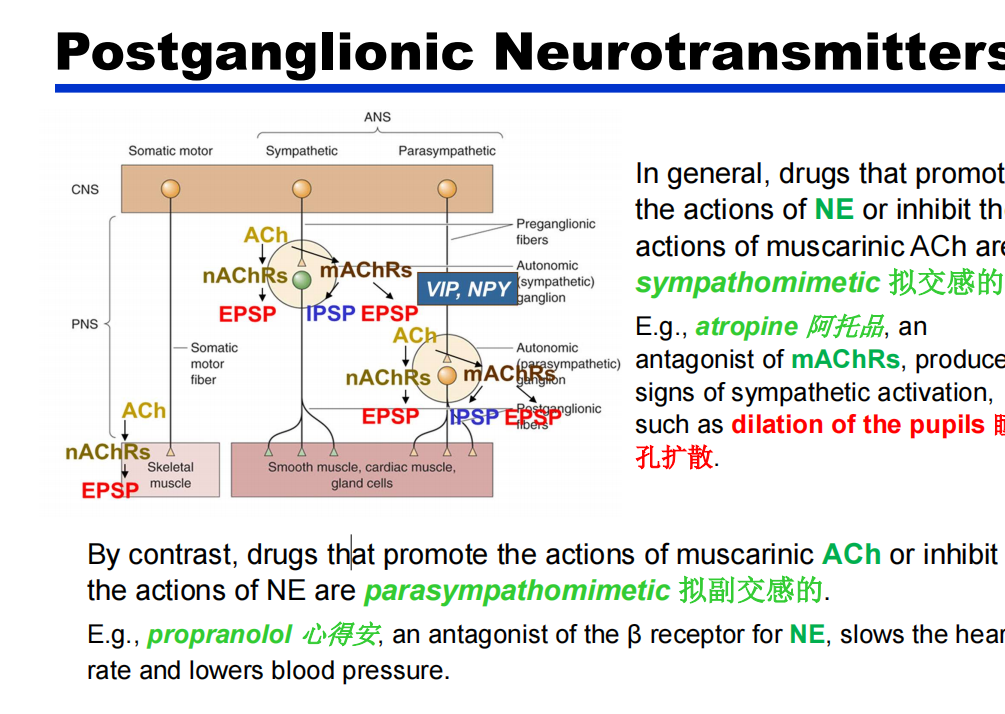
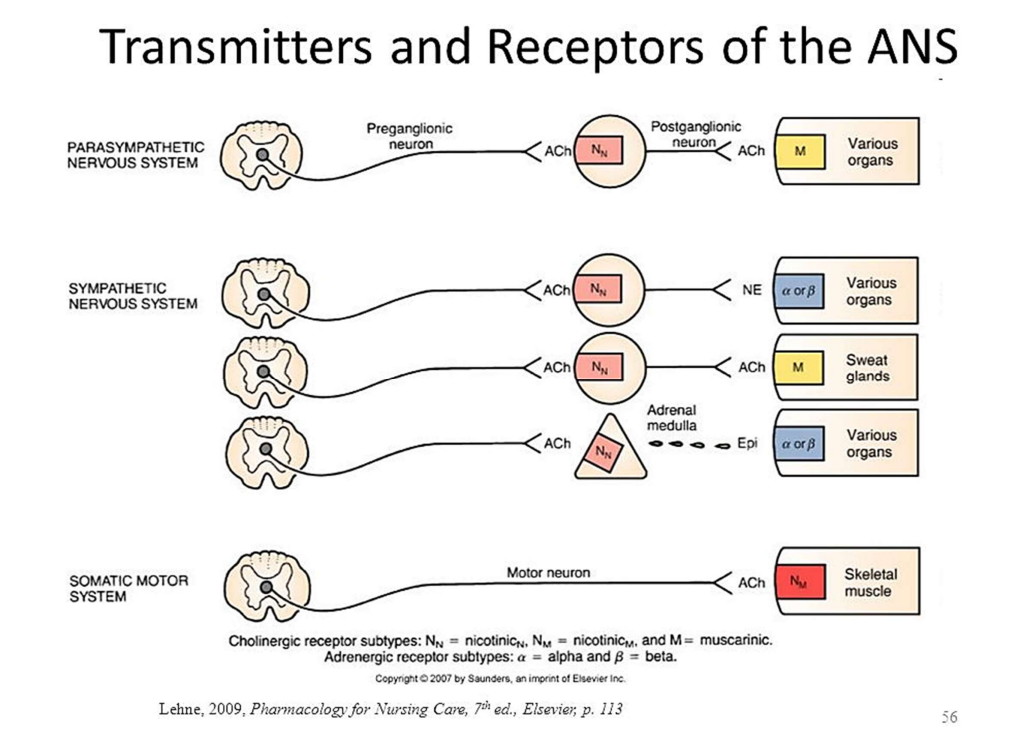 注:肾上腺髓质释放的ENE效果基本与NE相同,因此肾上腺髓质类似于被修饰的神经节。
注:肾上腺髓质释放的ENE效果基本与NE相同,因此肾上腺髓质类似于被修饰的神经节。 - The enteric division:可实现自我运行但离不开中枢神经系统!如下丘脑和孤束核NTS——PPT64-67
- 节前:除了Ach还有VIP、NPY(神经信号肽类—ppt42)
-
弥散神经系统:少数神经元、大部分源于脑干、大量分支、神经递质释放到细胞外液便于扩散
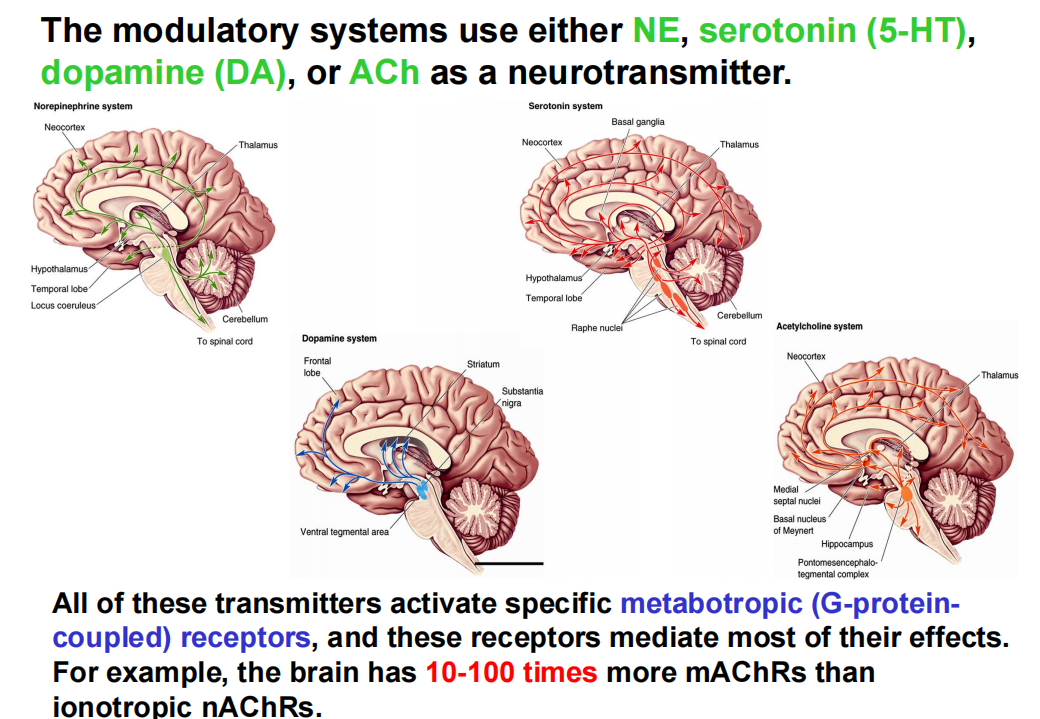


- NE system:蓝斑神经元的小簇投射出轴突,支配中枢神经系统的大片区域,包括脊髓、小脑、丘脑和大脑皮层。

- 5-HT(PPT79):中缝核沿着脑干的中线聚集,广泛地投射到中枢神经系统的各个水平。
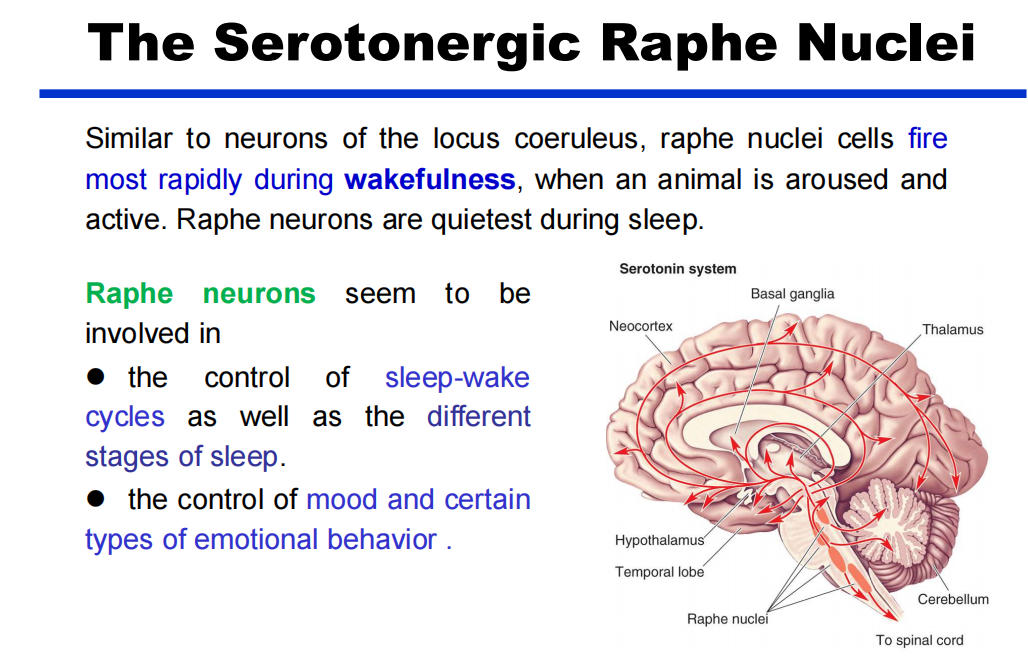
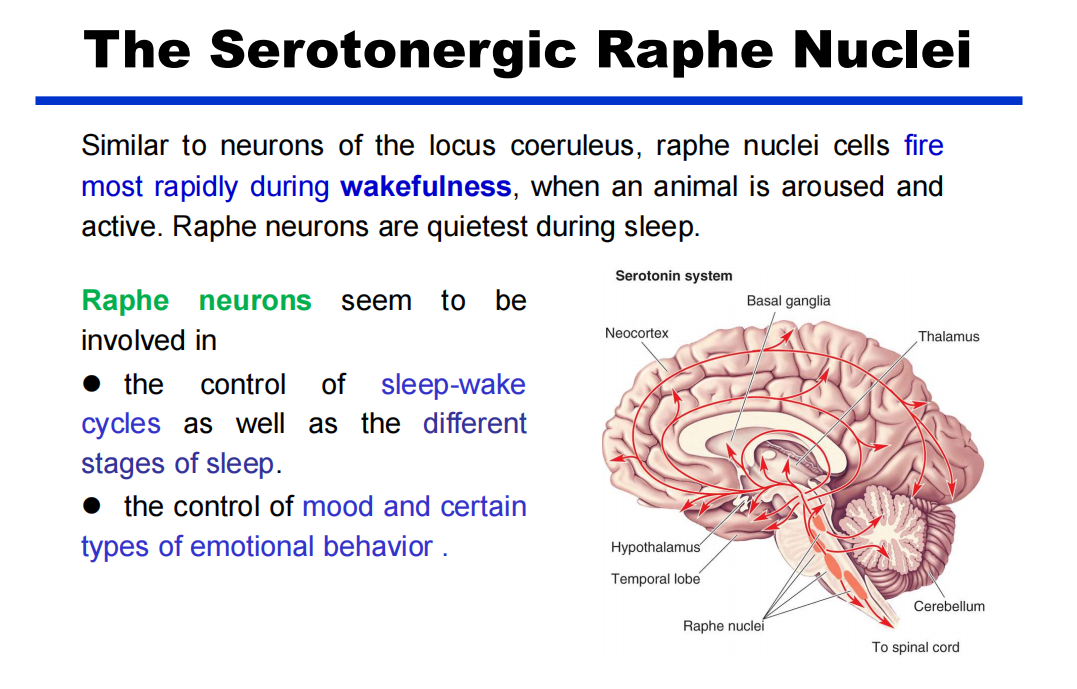 注:蓝斑和中缝核是上行网状激动系统的一部分,这意味着脑干的网状“核心”在唤醒和唤醒前脑的过程中。
注:蓝斑和中缝核是上行网状激动系统的一部分,这意味着脑干的网状“核心”在唤醒和唤醒前脑的过程中。 - DA:多巴胺能弥漫性调节系统来自黑质和腹侧被盖区。它们分别投射到纹状体(尾状核尾状核和壳核硬膜)和边缘和额叶皮质区。
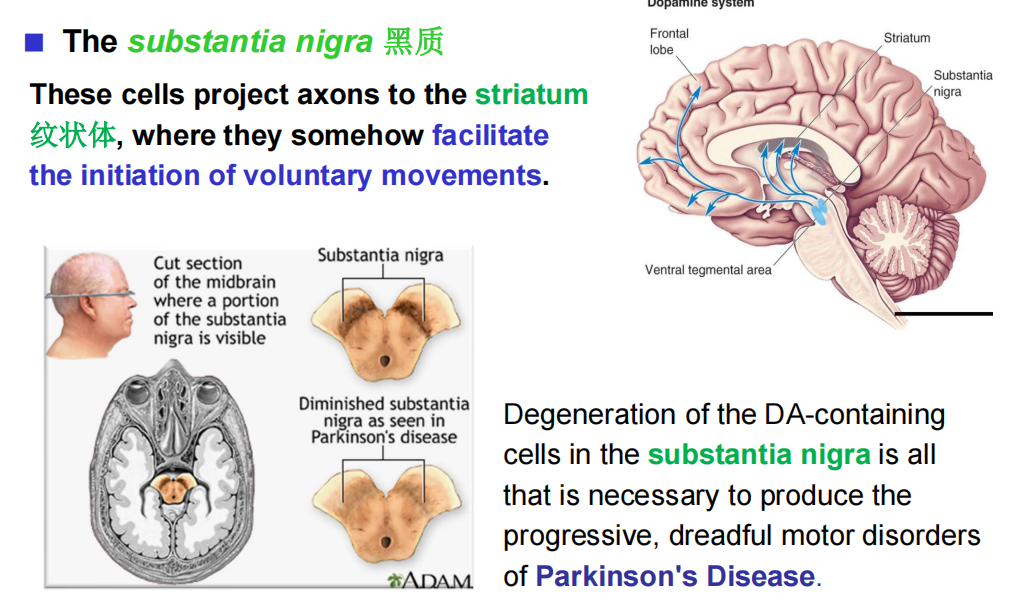 注:纹状体,帮助运动的起始
注:纹状体,帮助运动的起始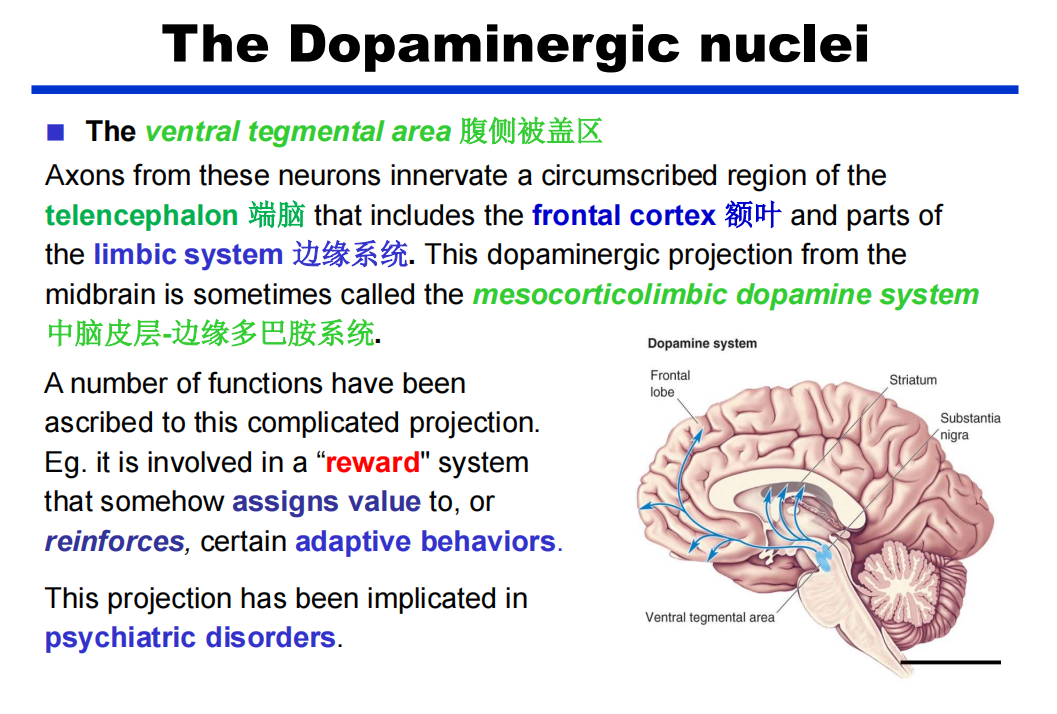 注:腹侧被盖区可以给事件的好坏赋值,帮助学习记忆的强化
注:腹侧被盖区可以给事件的好坏赋值,帮助学习记忆的强化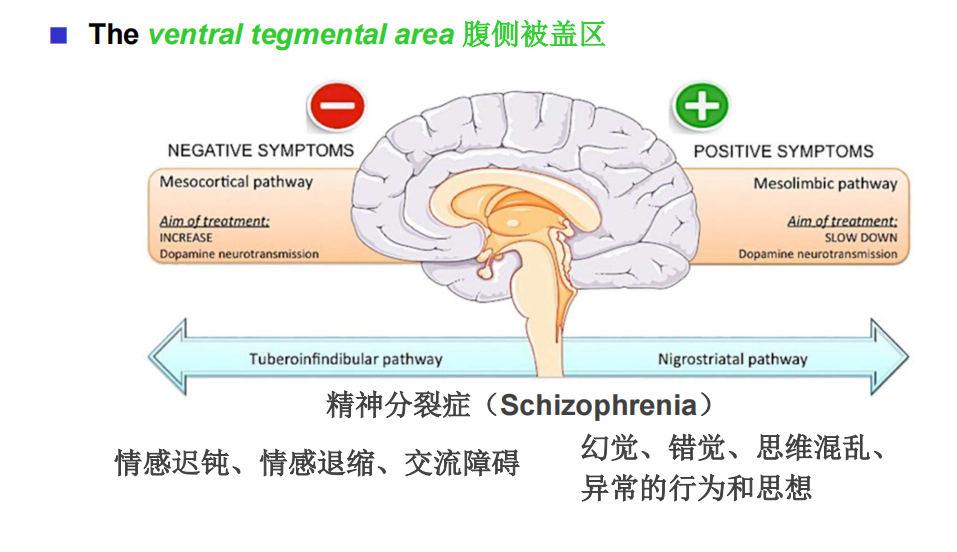
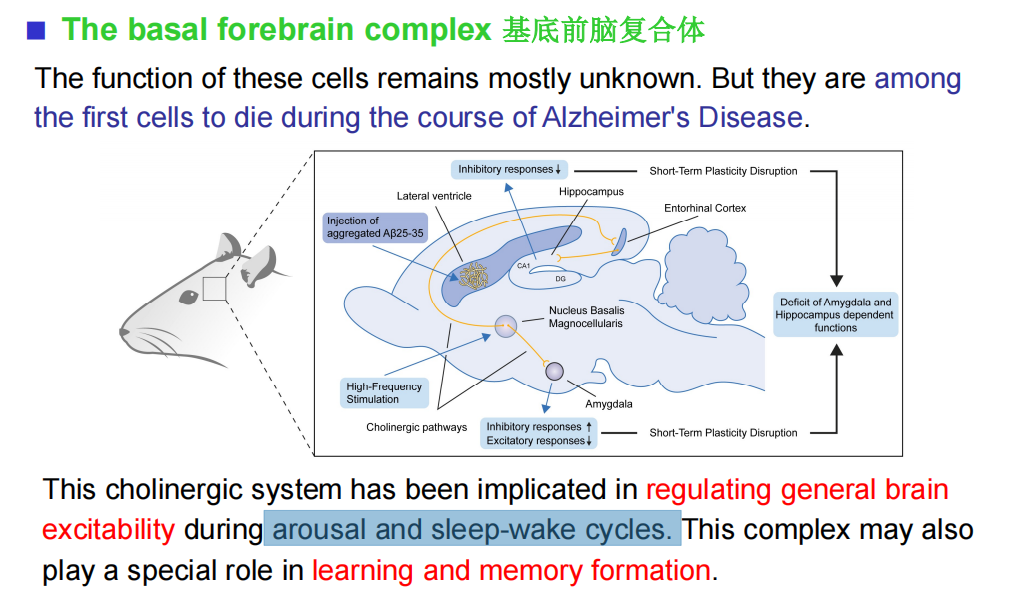
- ACh:阿尔茨海默有关
- 基底前脑:
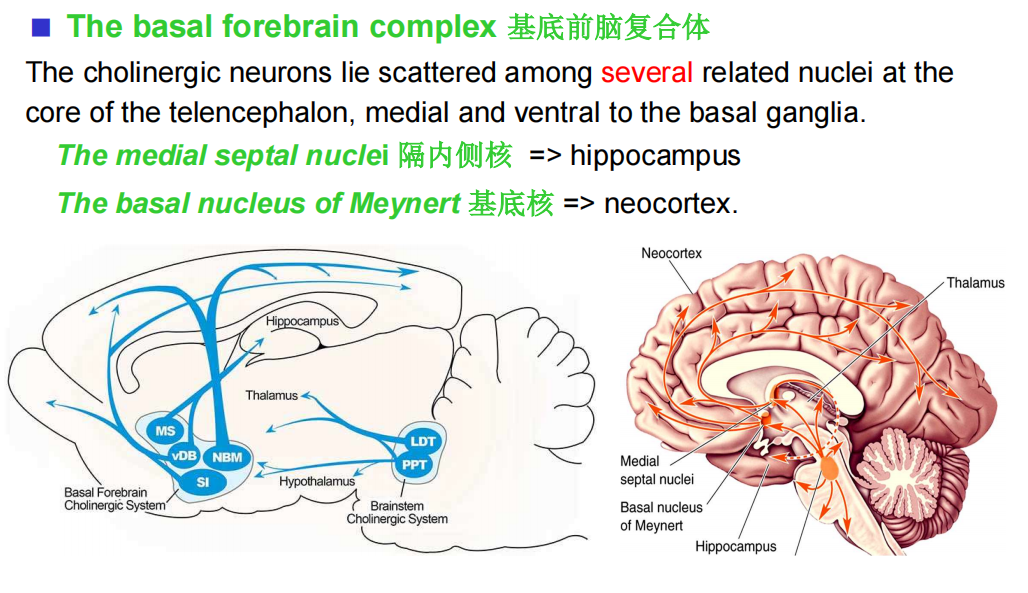
- 脑干:

- 基底前脑:
- NE system:蓝斑神经元的小簇投射出轴突,支配中枢神经系统的大片区域,包括脊髓、小脑、丘脑和大脑皮层。
-
Q
- 关于小分子神经递质和神经肽的区别,小分子神经递质在突触前末端合成,囊泡聚集在活性区(active zone);神经肽在胞体合成后通过囊泡运输到突触前末端,囊泡的分布较小分子递质远离活性区。为什么两者在神经元中的合成部位不同,其囊泡在突触前末端的分布位置也不同?

- 排尿过程的神经支配;紧张的时候想上厕所,以及吓尿了是怎么回事?
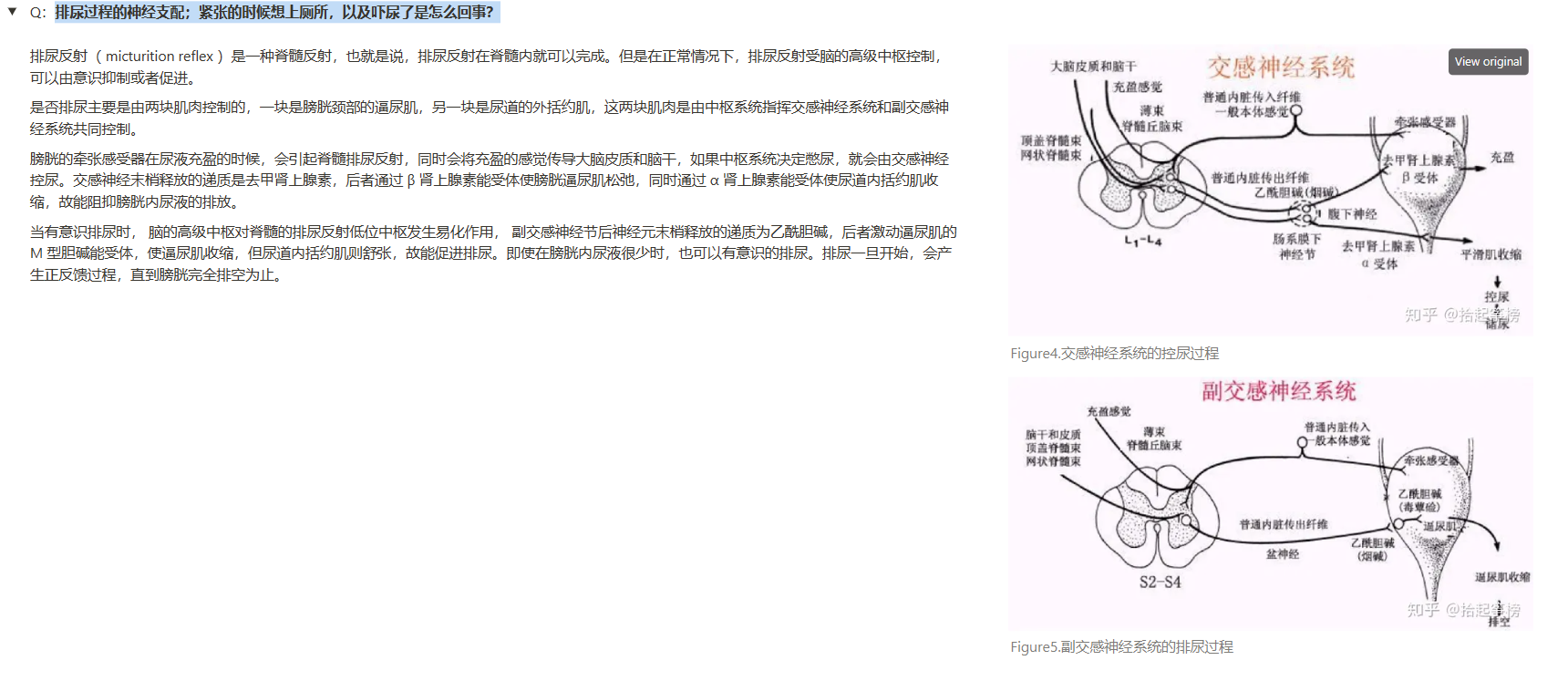

- 战场上受伤的士兵丧失了大量的血液后,常渴喝水,为什么?——答:当士兵失去大量血液时,肾脏分泌肾素进入血液。血液中的肾素增加促进血管紧张素II 的合成,血管紧张素 II兴奋下丘脑穹隆器的神经元。下丘脑穹隆器的神经元刺激丘脑外侧的细胞,引起强烈的口渴感觉
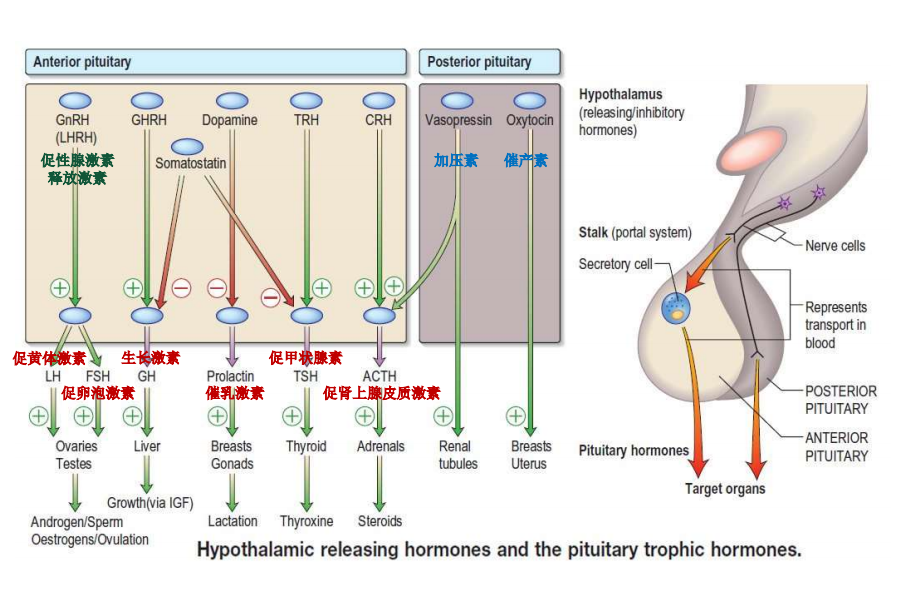
- 你整夜工作试图在限期内赶出一篇论文现在你正在疯打字,你一只文另一只眼睛盯着钟。详细描述下丘脑室周区怎样控制你机体对这种应急的生理反应?——答: 当人处于这种应急时,下丘脑室周区的小细胞性神经分泌细胞分泌促肾上腺皮质激素释放激素(CRH)进入下丘脑-垂体门脉循环。CRH 刺激垂体释放促肾上腺皮质激素(ACTH)进入体循环,从而促进肾上腺皮质释放皮质醇,皮质醇进入血液后,能作用于整个机体,动员能量储备,抑制免疫系统,准备来面对应急。
- 为什么肾上腺质通常被称为一个被修饰的交感神经节? 为什上皮质不被这样称? 答:当人处于应急或时,交感神经兴奋,通过兴奋交感神经节前纤维刺激肾上腺素髓质分泌肾上腺素和去甲肾上腺素,这些激素对靶组织的作用与交感神经兴奋引起的作用几乎相同,所以说肾上腺髓质是一个被修饰的交感神经节。然而肾上腺皮质,是在小细胞性神经分泌细胞释放促肾上腺皮质激素释放激素(CRH),通过门脉循环作用到垂体,刺激释放肾上腺皮质激素(ACTH)然后ACTH 通过体循环刺激上腺皮质释放皮质醇。虽然最后引起的作用和交感神经兴奋有些是相同的,但是整个过程没有涉及到交感神经节前神经元,所以不能被称为一个被修饰的交感神经节
- 很多著名运动员和演员由于服用大量可卡因而突然死,通常死亡的原因是心脏竭。你是如何解释可卡因的这种外周作用的?——答:可卡因主要阻断多巴胺的重摄取,导致其在轴突间隙内的时间延长,从而延长和加强多巴胺的作用,能够增加心率和血压,达到兴奋效应。当服用大量可卡因时,导致有大量多巴胺聚集在突触间隙,使心肌细胞过度兴奋超出负荷而出现心脏衰竭
- 脑内的散性调节系统与点对点的突联系系统之间有什么区别? 列出四点区别——答:1) 点对点的突触联系是短时程的,而弥散性调节系统是长时程的。 2) 点对点的突触联系是短距离的,而弥散性调节系统是长距离的。 3)点对点的突触联系每个神经元与很少的突触后神经元接触,而弥散性调节系统的神经元与100K以上的突触后神经元接触。 4)点对点的突触联系释放的递质仅局限于突触间隙周围,而弥散性调节系统的递质能扩散到很多神经元。 5)点对点的突触联系在CNS 和 PNS 都有分布,而散性调节系统存在于CNS
- 在哪些行为状态下的去甲上腺素能神活动? 自神系统的去甲肾上能神经元活动?——答:环境中新的、非预期的、非疼痛的感觉刺激最容易引起蓝斑神经元的兴奋,当运动、焦虑、情绪激动时会引起去甲肾上腺素能神经元兴奋
- 为什么男生没有例假?——男女的Hypothalamus-Pituitary-Gonads存在明显的雌雄差异,在雄性的HPG系统中,受到刺激的是睾丸,睾丸分泌的睾丸激素作用于丘脑和垂体是一个直接的短暂迅速的负反馈,所以男生的分泌可能只会有一个短暂的daily的周期。而对于雌性来说,卵巢在受到激素刺激后分泌的雌二醇和黄体激素与丘脑和垂体间既存在有负反馈,还有正反馈,这两种反馈的作用结果就是激素的水平在一个较长的时间内会有一个缓慢的上升变化过程,最终就导致了女性的一个漫长的生理周期。
- 肠道菌群影响HPA轴(下丘脑-垂体-肾上腺轴)的机制有哪些?——肠道的神经系统很复杂,但是大体上是相对独立的。肠道系统常常不报告中枢神经系统,而自己做出决策,比如你不能自主的控制什么时候想要排便等。但是肠道系统毕竟属于身体的一部分,肠作为一个外周的器官也是存在有上行的突触到中央神经系统的,(或是其他什么原因),因而也不是完全独立的,比如你可以感受到肠胃的疼痛。因此肠道菌可能就是通过影响肠道神经系统进而去影响这些中枢神经系统。(先简单介绍一下HPA轴)(可以再介绍一下肠道菌群与肠道神经系统的关系)
- 鸡性转也是因为垂体吗?——垂体是性表征的器官,不是决定性别的器官。将公鸡和母鸡的垂体交换后并不影响其功能,也不影响鸡的性征和表型。决定性别的首先是基因,然后再是激素。先说明一下性转是由雌转雄还是雄转雌,还是都可以;最后可以简单解释一下基因是如何影响性转的,因为这里可能会有“激素的合成依赖于基因”这样的误解
- 可以把非本能行为变成本能行为吗?——可以说既能又不能,在一个个体的水平上来说,你可以把一个非本能行为变成本能行为,比如我们聊天中说“这就像我们的第二本能一样”,这个动作你重复成百上千次后就已经不需要大脑去思考了,相当于一个habit,在经过长期的训练后变成了一个固化的运动环路,所以从个体的水平来说,你可以把一个非本能行为变成一个很快速的不需要经过思考的类似的本能行为;但是从人类群体的进化角度讲,这种通过训练获得的个人水平上的本能是不能通过群体传递给下一代的。
- 能将个体不放在群体而是单独培养来研究本能吗?——本能行为就是不需要他人授予,与生俱来的一个能力,这在蜥蜴和鸟等动物上最容易证明,因为这些动物不是哺乳动物,哺乳动物的行为很难界定是不是本能行为,因为在他们的成长过程中总需要母亲的喂养,有其他个体的一个介入,但是我们将一个鸡蛋拿过来单独孵化,在从未见过其他个体的情况下,鸡仍然能表现出一系列很复杂的很像它的同类在野生条件下的行为,比如筑巢、迁徙等。
- 狼孩不训练也能直立行走这算本能行为吗?——首先狼孩是不能学会直立行走的,他只会和狼一样四肢着地爬着走。在哺乳动物里其实已经很难去界定一个具体的本能行为,因为自出生起动物就开始和其它个体一起生活,导致它的行为总是会受到各种环境经验所调控的,那么这时候本能更多的可能是作为一个背景,最后的行为总是一个基于此背景的习得性的结果。因为狼孩是被狼养的,如果被其他动物养就可能不会具有像狼那样的生活习性,所以狼孩爬着走行为并不是一种本能,而是基于他本身所具有的运动能力,去模仿狼的行为的一个习得性结果。但是他会走这件事情,或者说运动能力可能就算是他的本能。本能行为在哺乳动物中肯定是有的,但是他总是和习得性行为混合在一起,所以我们最后看到的就不是单纯的本能或习得性行为,而是两者的混合,有时候是本能多一些有时候习得性更多。
- 皮质醇会杀人吗?——皮质醇作用的一个后果:神经元通过电压门控离子通道吸收更多Ca2+。慢性剂量的应激导致神经元钙超载,并导致神经元死亡(兴奋性中毒)。每天注射皮质酮(大鼠皮质醇)数周,导致许多皮质酮受体神经元的树突萎缩。几周后,这些细胞开始死亡。 也发现了类似的结果,当每天给老鼠注射激素而不是每天给老鼠注射激素时。 狒狒体内:无法逃脱笼子里的“顶头狒狒”,许多下级雄性狒狒死于胃溃疡、结肠炎、肾上腺肿大和海马神经元的广泛退化。这些皮质醇和压力的影响类似于大脑老化的影响。 在人类中,接触极端暴力可能导致创伤后应激障碍(PTSD),症状包括高度焦虑、记忆障碍、侵入性思维、大脑(尤其是海马体)的退行性变化。
- 关于小分子神经递质和神经肽的区别,小分子神经递质在突触前末端合成,囊泡聚集在活性区(active zone);神经肽在胞体合成后通过囊泡运输到突触前末端,囊泡的分布较小分子递质远离活性区。为什么两者在神经元中的合成部位不同,其囊泡在突触前末端的分布位置也不同?
脑和行为的化学控制——下丘脑与自主行为(思维导图见上)
- 下丘脑的室旁区——调控功能很重要
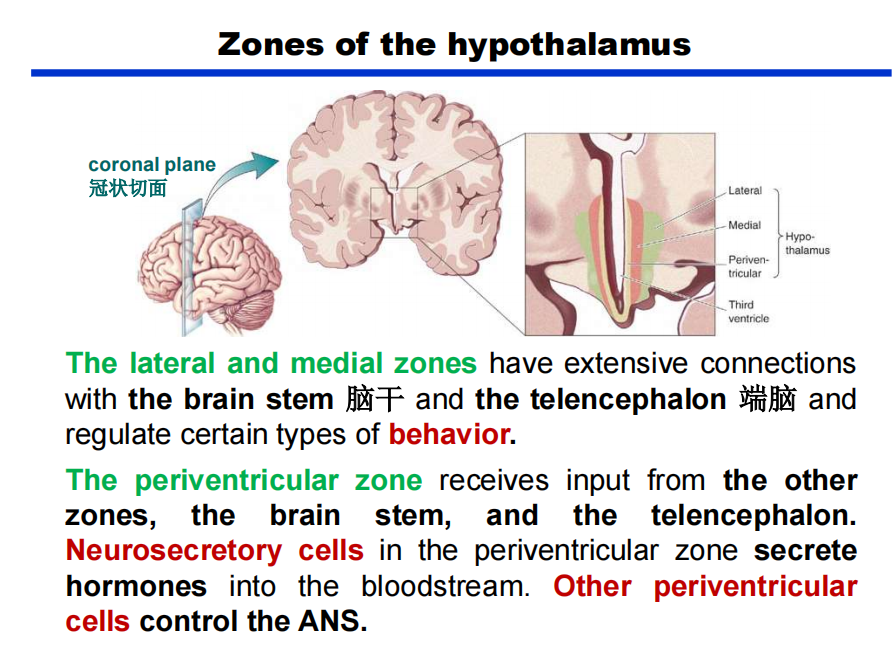

- 关于室旁区第三条的叙述:
- 大细胞神经分泌细胞是下丘脑神经分泌细胞中最大的一种,它在视交叉周围延伸轴突,沿垂体柄向下,进入后叶。
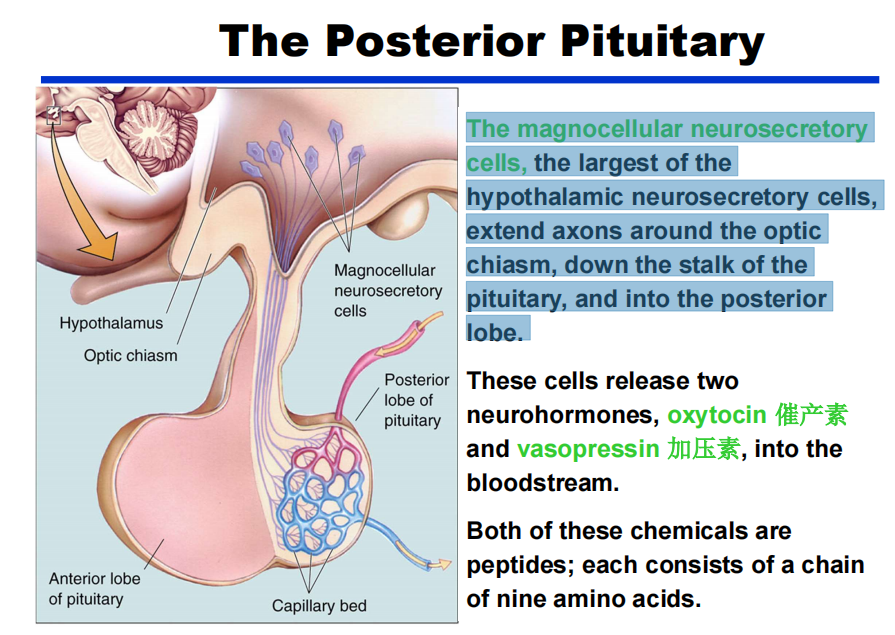 注:如抗利尿激素:
注:如抗利尿激素: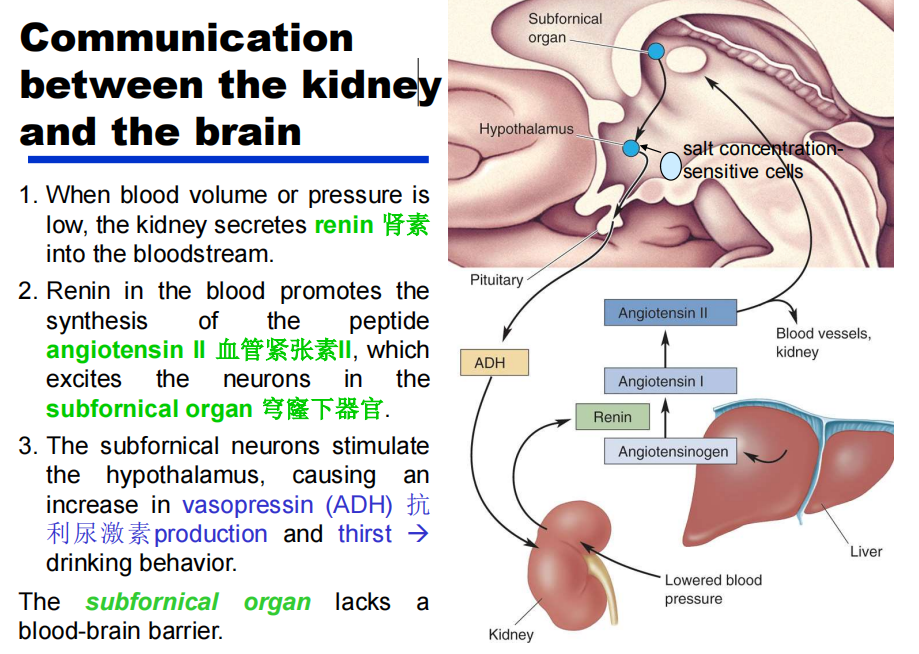
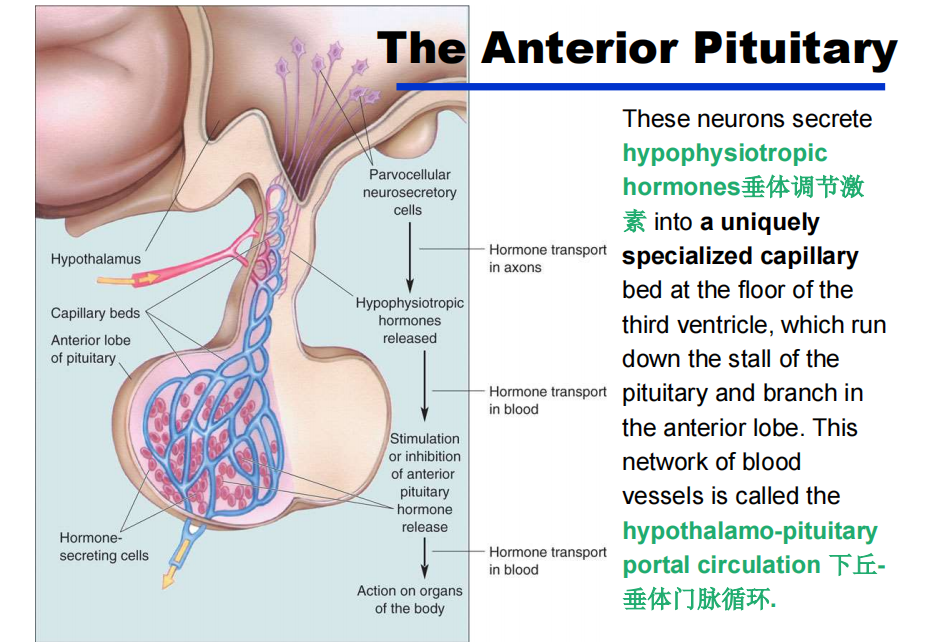
- 小细胞性:脑室周围区域的微小细胞小细胞性neurosecretory细胞不会将轴突一直延伸到前叶;相反,它们通过血液与目标进行交流。这些神经元分泌垂体调节激素到第三脑室底部的一个独特的特殊毛细血管床,该床沿着垂体和前叶的分支运行。这个血管网络被称为下丘-垂体门脉循环。
- 大细胞神经分泌细胞是下丘脑神经分泌细胞中最大的一种,它在视交叉周围延伸轴突,沿垂体柄向下,进入后叶。
- 关于室旁区第三条的叙述:
- 关于稳态:
- 下丘脑与瘦素:
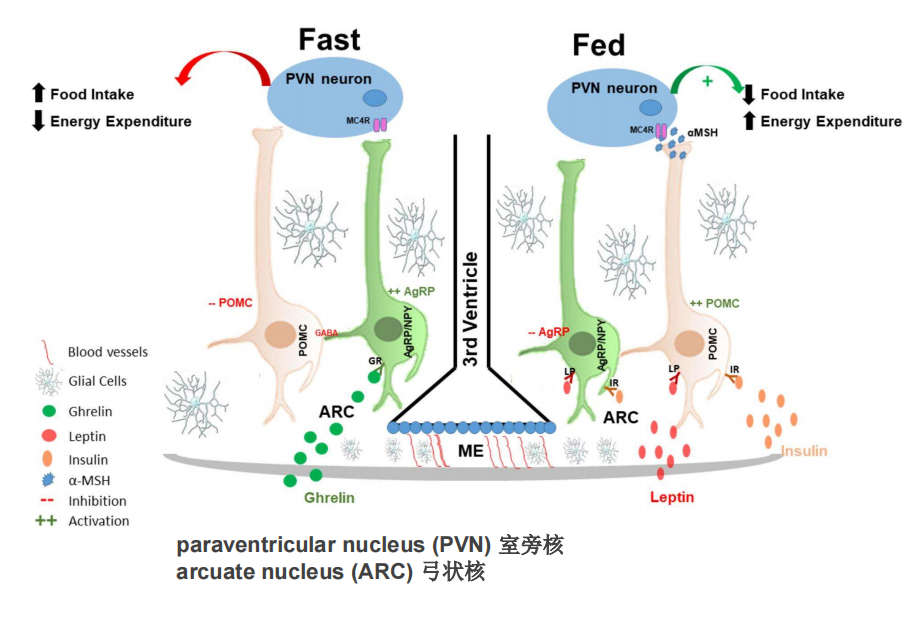
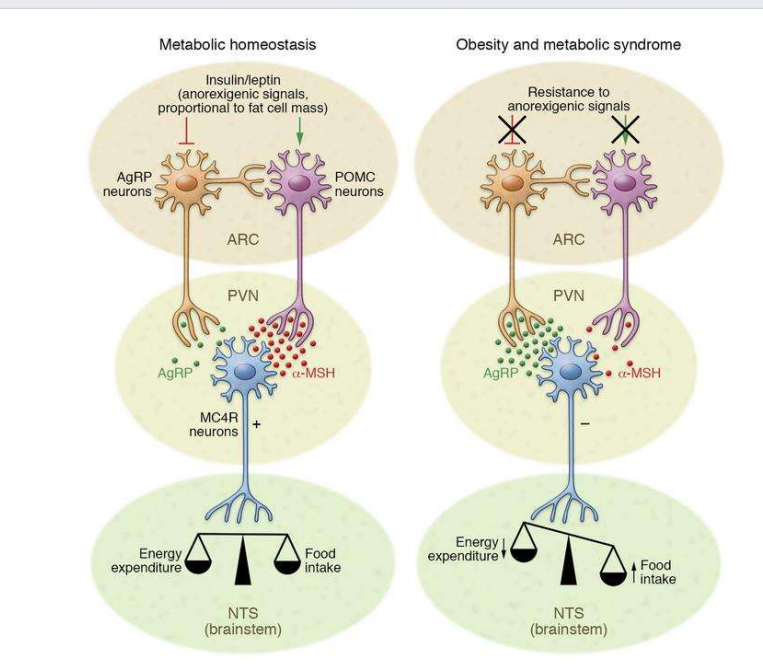 注:抑制与饥饿的不足会产生肥胖
注:抑制与饥饿的不足会产生肥胖 - 视交叉上核(SCN):与节律的关系——体外培养实验中细胞间的相互作用增加了转录-翻译反馈回路(TTFLs)的稳定性、一致性和振幅,并使单个时钟细胞同步,导致精确组织的基因表达和神经活动节律。
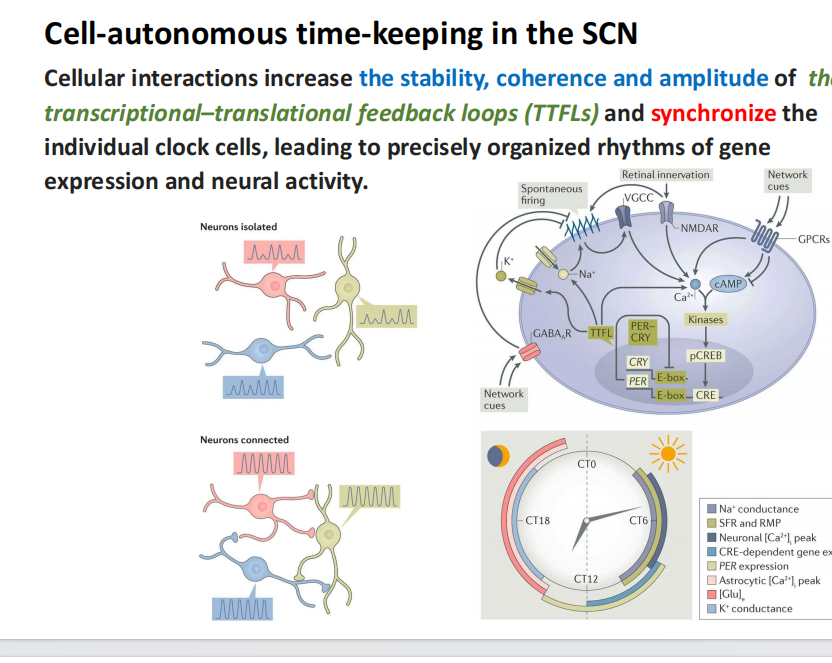
- 压力与CRH、ACTH——皮质醇(能量储存与免疫系统的抑制)
- 下丘脑与瘦素:
- 本能行为——果蝇的启示(跨器官的研究——开放式神经系统+denormal规律的发现——启示对人脑的研究+节律研究+认知功能研究)
学习和记忆的环路机制
- 海马(hippocampus,通路见PPT18)对于获得新的外显记忆(可陈述)具有重要作用。前额皮质对于工作记忆具有重要作用,而小脑和基底神经节则参与多种运动学习过程。记忆的提取不需要海马
- The circuit organization of the mammalian hippocampus(哺乳动物海马体的回路组织)
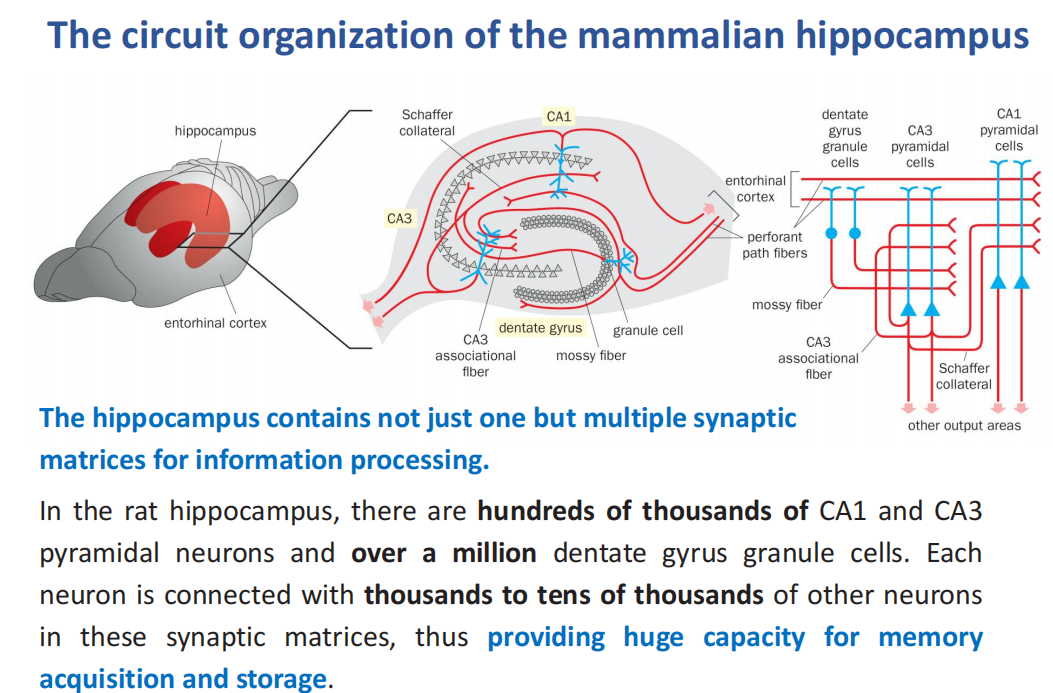 注:在大鼠海马中,有成千上万的CA1和CA3锥体神经元和超过100万个齿状回颗粒细胞。每个神经元都与这些突触矩阵中成千上万的其他神经元相连,从而为记忆的获取和存储提供了巨大的容量。
注:在大鼠海马中,有成千上万的CA1和CA3锥体神经元和超过100万个齿状回颗粒细胞。每个神经元都与这些突触矩阵中成千上万的其他神经元相连,从而为记忆的获取和存储提供了巨大的容量。 - 希波理论:当细胞A的轴突足够近,足以刺激B细胞,并反复或持续地参与激发它时,一个或两个细胞发生一些生长过程或代谢变化,使A的一个激活B细胞的效率提高
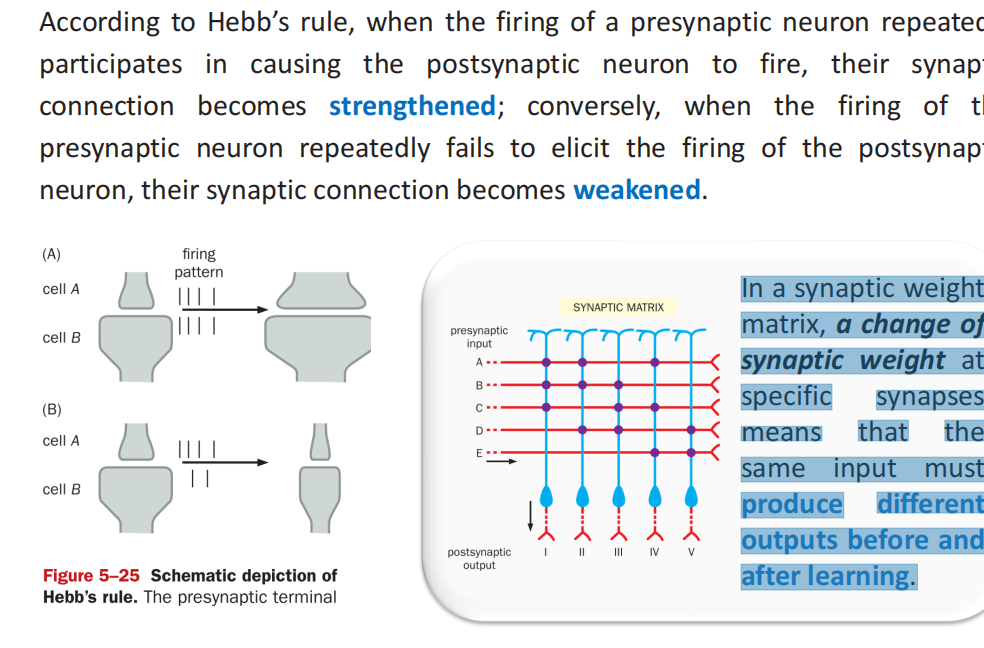
- 突触的可塑性:学习记忆的关键——PPT22-31开始
- 海兔习惯化和敏感化的过程:涉及cAMP信号通路
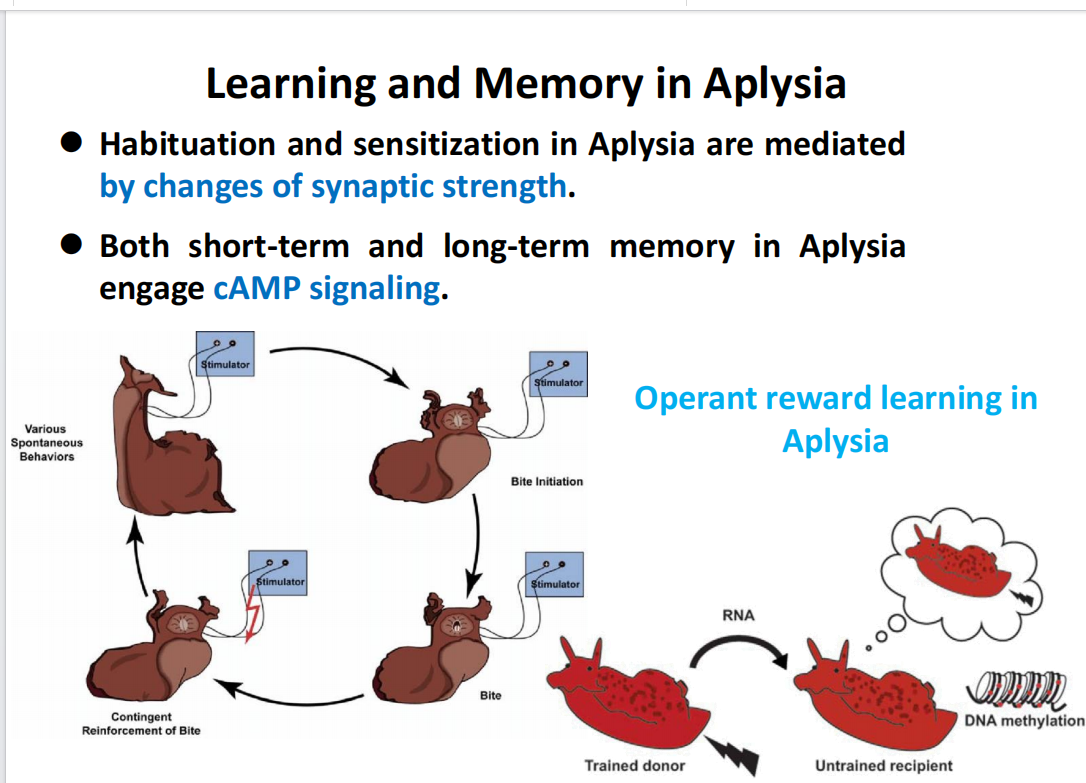
- 线虫在学习和记忆中的研究——筛选记忆弱的突变体
- LTP不依赖海马
电与突触——见孙老师资料
Q:
- 突触后膜离子型受体是如何关闭的?——离子型受体即配体门控的离子通道,当有配体结合时通道就会打开,当相应配体离开,通道关闭。所以这个问题在于突触间隙的小分子神经递质是如何被清除的。突触间隙的小分子神经递质有多种清除方式:被动扩散,被突触前膜回收,被周围的星型胶质细胞社区以及被相应的酶降解。
- 如何正确理解兴奋性突触后电位(EPSP)和抑制性突触后电位(IPSP)?
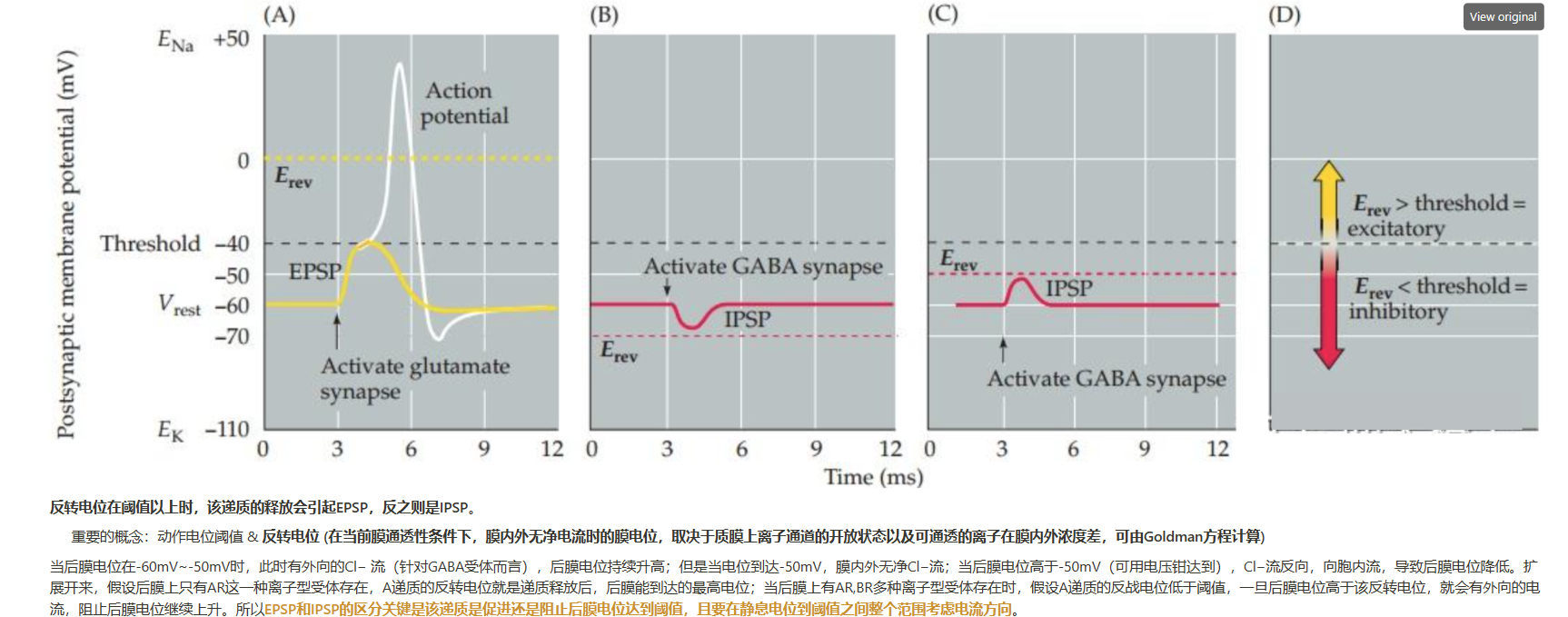
- 如何理解GABA在发育中的大脑是兴奋性递质,但在成熟大脑中却是抑制性递质?——递质释放对突触后膜达到阈值是有利还是不利取决于反转电位的高低,而反转电位则取决于膜通透性(离子通道的开放)以及膜内外的离子浓度差。GABA的离子型受体是Cl−通道,发育中和成熟的大脑胞内外Cl−差不同。发育中的神经元膜内外的Cl−浓度主要由 Na+−K+−Cl− 协同转运蛋白(NKCC1)决定,其将Cl−泵入细胞,因此胞内Cl−浓度高于胞外,这时的反转电位高于阈值,GABA的释放可以引起突触后膜发放动作电位。成熟的神经元膜上主要表达K+−Cl−协同转运蛋白(KCC2),其将Cl−泵出细胞,因此胞内Cl−浓度低于胞外,GABA会造成突触后膜超极化。所以,递质在突触后膜上的作用效果是由递质、相应受体、相应离子的膜内外浓度差三者共同决定的。此外,GABA在神经发育过程中的兴奋性作用的意义尚不明确。
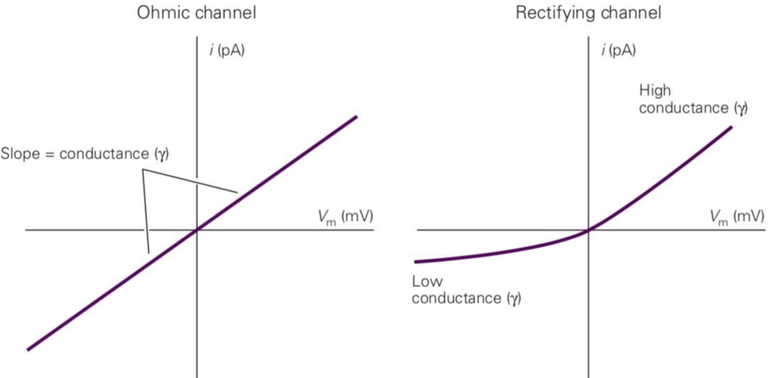
- 内向整流钾离子通道是如何工作的?
——首先我们需要理解“整流”的概念。许多离子通道开放程度与电压无关,其伏安特性曲线如图 2中左侧所示为线性的:随着电压的升高,电流也升高,斜率即为电导。但有一些通道的离子开放程度是电压依赖的,其伏安特性曲线则是非线性的,在图2右侧的例子中:电压较低时,电流基本不随电压的变化而变化,而在电压较高时,二者又呈线性关系。
内向整流钾离子通道在神经元静息状态下正常开放,钾离子可以外流。在去极化状态时,电压升高,通道被Mg2+ 等离子堵住而关闭, 钾离子外流被阻断。
我们可以这样理解为什么去极化时要阻止钾离子外流:去极化时钠离子内流使得膜内正电荷增多,有利于形成动作电位,这时如果钾离子外流也增多的话会损失膜内正电荷,这不利于形成动作电位,所以内向整流钾离子通道此时阻止钾离子外流。
- 静息电位和动作电位中钾离子的跨膜渠道相同吗? ——不相同。 静息状态:K+通过泄漏通道流向胞外 动作电位:K+主要通过电压门控K+通道流向胞外 离子泵在两种状态下均工作,将K+泵向胞内,Na+泵向胞外 关于内向整流K+通道的部分详见“Q4、内向整流钾离子通道是如何工作的?
- 如何利用电压钳研究动作电位?——神经元动作电位的电压是连续变化的,而电压钳可以将膜电位钳制在某一特定数值,从而研究神经元动在该特定电压下的工作机制。如图所示,电压钳将膜电位钳制在-26、0、26、 52 和 65mV,根据记录到的补偿电流,我们就可以得知在此电压下的神经元的工作状态。
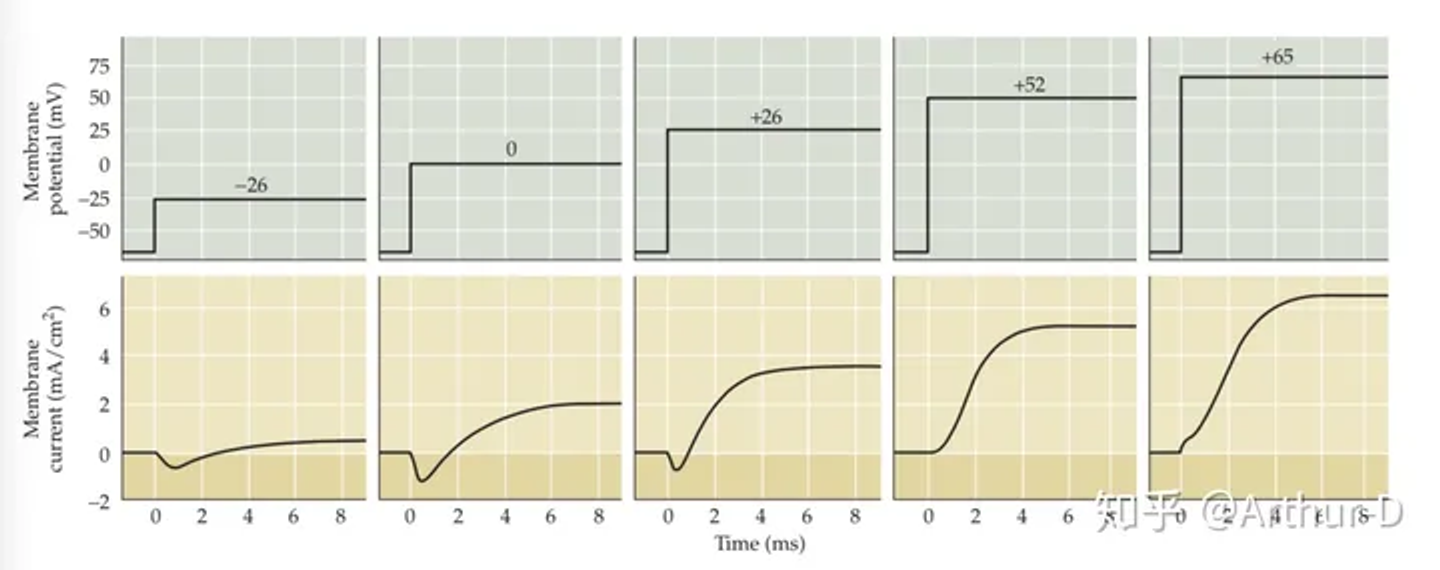
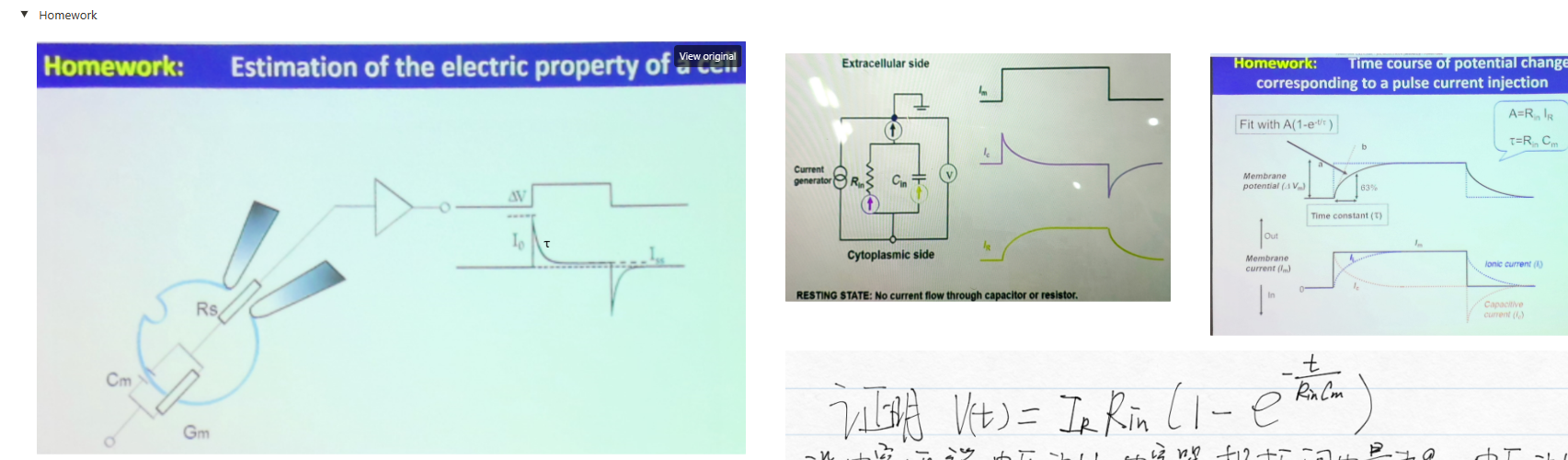
- 作业: