神经元的病毒标记与示踪
背景
灰质:胞体;白质:髓鞘化的纤维——越高等白质越多【axon增加的比例越多——智慧的象征】
高尔基染色:利用神经元的嗜银性质
网状学说:Reticular Theory【神经系统完全连通——高尔基】
突触学说:卡哈尔
不同尺度的脑联结图谱:纳米【电镜扫描】——微米【病毒示踪】——毫米【核磁】限速步骤在于轴突总长度
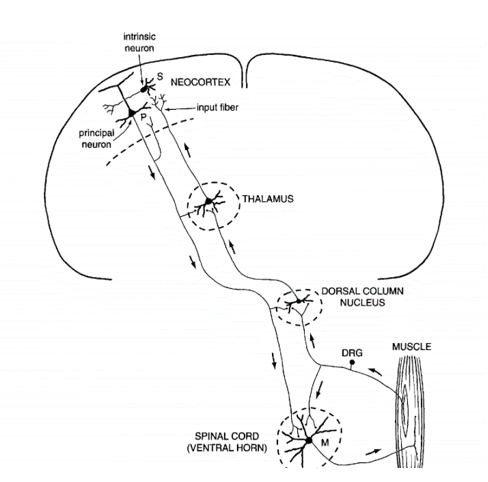
主要负责思想的细胞 :PC细胞【锥体细胞—兴奋性细胞,各个层都存在(不同层的靶区不一样),主要在第五层】,胞体大,且有很长的树突,往下的轴突可长达1m,延展到脊椎,控制我们手的运动等。——终极目标,阐明不同神经元之间的联系 【可从形态来猜测神经元之间的连接关系和功能,比如锥体细胞为什么需要那么多树突——搜集信号能力强,对大脑中的很多功能起绝对作用】人为什么不能自己给自己挠痒痒?——锥体细胞轴突的分支到丘脑,丘脑对皮层柱进行反馈【整合信息,告诉大脑我的手移出去——无surprise】
突触前神经元引起的突触后电位不会立马兴奋或抑制神经元,他只是产生了突触后电位
锥体神经网络上游的输入并不是等权的【计算机神经网络是等权的】——其计算能力非常强,强于多个二极管。
大脑中长程投射很少,大部分是local——这也突出了长程投射的重要性
脑联结图谱的重要性 :
神经元的稀疏标记 :一开始是随机标记,后面发展到特异性的稀疏标记
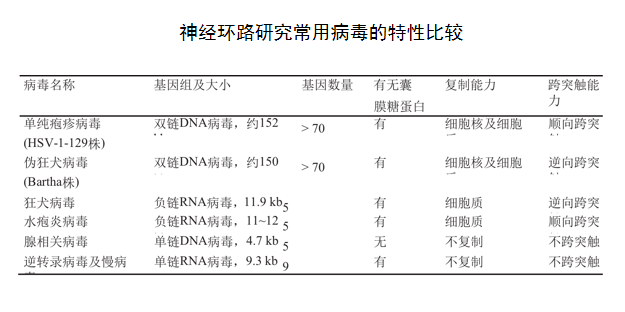
嗜神经病毒 ——如腺相关病毒【不复制、不跨突触(安全)、载量小单足够】
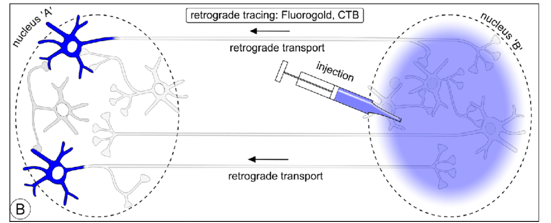
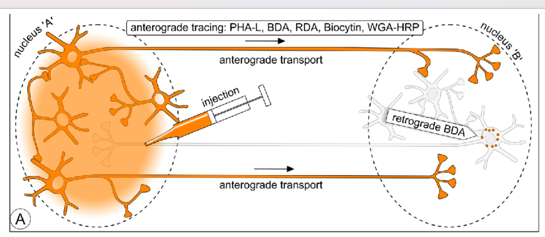
逆行标记:如CTB【霍乱毒素B亚单位】,RV【狂犬病毒,看单细胞的上游】,逆向病毒示踪,打在末梢,点亮胞体
顺行标记:如HSV【单纯疱疹病毒】,打在胞体,点亮末梢
AAV【 长程的circuits,因为绝大部分不跨突触 ,只标了纤维和末梢,看投射】:2、9标记神经元
不跨突触:
跨单突触
AAV1-cre 【顺向】、H129▲TK-tdT、Hs06逆行:CVS-N2c、RV【狂犬病毒】
跨多突触
WGA【同学讲解】:可正向逆向跨突触示踪?——mWGA只可以单突触顺行标记【微环路】
鼠系 :AI系列【含Loxp元件】
脑联结图谱 :【连接组:把神经元尽可能分解成不同的类型,然后看某一类型的结构联结——可结合光镜+电镜进行重构】
连接组重构目标是:广泛到全脑【fMRI+DTI】、精细到突触【fmost介观】、区分细胞类型、动态重构
Frontal cortex:轴突的连接最长最密集——与意识有关?
胼胝体:左右侧神经元的镜像投射
连接状态:兴奋型——Spiny(比较粗)轴突;抑制型——smooth轴突(细)形成突触
决定神经元和神经元之间的连接(是否建立突起/突触联结):通过位置?越近概率可能越高?;两个神经元同时放电 ,他们建立的所有突触会被增强【可塑性】——【电镜水平】通过突起大小(还有囊泡的数量等)判断强弱;金标准 ——A神经元被刺激,B神经元产生的反应【膜片钳Patch两个神经元】
神经元分类:同一类型的神经元有相似的基因表达【还原论策略】
形态学
电生理性质
基因表达——同一个基因代表的细胞也有很多亚型,包括形态也不同——难以分类
脑的连接矩阵:topography——所有不同位置皮层的神经元,会靶向同一个区域——大脑结构存在很强的规律性【无特异性、上游细胞的表型未知——用cre系来优化图谱】
脑联结图谱的构建——透明化
关于透明化BB—BED:
Thy1品系:神经元特异—看清楚axon、胞体等细节说明透明化效果好
Ai140 鼠【来源于ailand研究所】:存在一个自放大过程 CreERT2 (雌激素受体依赖,打入Tamoxifen来刺激Cre的表达,根据T量的多少实现稀疏标记) 鼠系交配得到杂交后代,这个后代发育过程中并不会诱发Cre的表达,避免发育的干扰。拓扑关系:如手指不同感受野【感觉神经元】到脊髓的映射始终分离
关于神经元的差异:内在基因、基因相同可能形态结构【诱导因子诱导他们朝某一方向】也不相同【没有完全相同的两个神经元 ,如在发育的过程中相同基因类型的神经元会受不同小分子信号梯度的诱导导致不同神经元向不同方向延伸,即形成生长锥 】,这时,可以通过看他们投射的范围来判断类别、发育生物学的角度【对相同基因表达的谱系进行示踪】
小鼠大脑透明化:大部分情况不需要切割就可以进行成像了。
VISoR2:
包埋:水凝胶包埋【形变少】
切片:先切片,再透明化【250个 300um】
透明化:Puclear取代传统的SDS+RIMS
成像:面成像【斜着扫描】+样品移动【S形】
丘脑到前额叶皮质的投射——在IV层
柱状深层结构 :浅层进深层出(信息流)相关性并不能代表真实神经元之间的关系,得不到因果关系 ——如中间神经元的存在【抑制另一个神经元产生pulse,抑制神经元是方波】;如神经元钙活动相似——但可能他们的上游不同
LASSO【核磁】:去除冗余,使不等于0的β的数目最小【让冗余的神经元数目最小】
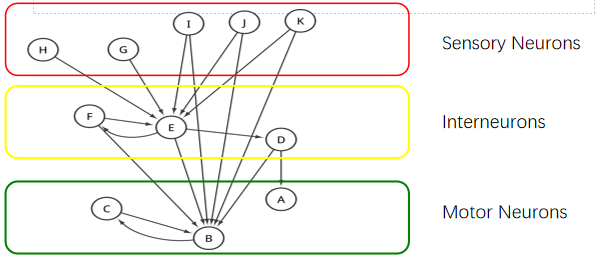
Random Surfer Model:评估神经元的重要性【信息流】——发放(sensory)+整合(Interneuron)+运动执行(motor neuron)——越高等的动物在此基础上加了很多Feed back【反馈】;并非输入最多的神经元就是最重要的神经元【如C 和B】,如果一个神经元和很多输入的神经元建立联结,那么它也很重要【相当于牛顿引用了你的文献】
网络的构建——长程+短程
透明化技术:
脑图谱构建之fmost:
单神经元分析技术
稀疏标记:【皮层下神经元轴突是由树突发出】
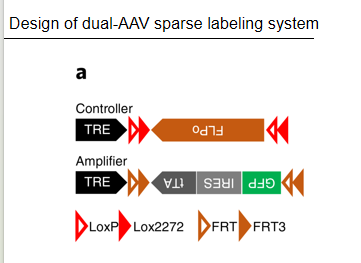
Flpo【还有ERT2-Cre】:【结合Ai140鼠系使用】Cre减去Flpo—Flpo切FRT【放出荧光】——二级自放大【只标记Cre阳性,1w个也较稀疏】
Ai140
控制病毒注射位点标记【AVV-Retro-Cre:打末梢,点亮胞体(胞体含cre才亮)】
投射谱研究的目的与意义:用Axon projection【靶区决定:可能是多个靶区】定义出的神经元是最好的——直接和功能相关【单个基因往往从功能角度难区分开来】;聚类——区分不同的投射模式(转录组水平亦可分开)【IT投射到2、3层,PT投射到低5层【第五层的椎体神经元传入到脊髓——burst 会帮助信号传递更远】,CT投射到第6层(主干保守)——看几百个和看几万个也是n个类别】;功能+发育轨迹的分析
如何衡量两个神经元【两棵树】的差异 :两个曲线分解成三角形【通过节点分解】——三角形面积之和,面积越小,距离越近随着胞体位置梯度的下降,会改变投射区域(靶区重心再移动)【类似于视网膜向上丘投射,约medial的,投的越anterior;越Lateral的地方,投的越postieral】
总结:
没有两个神经元是相同的 - 单神经元投射组的巨大变异性。
只有少量的单神经元重建不足以产生重要的见解。
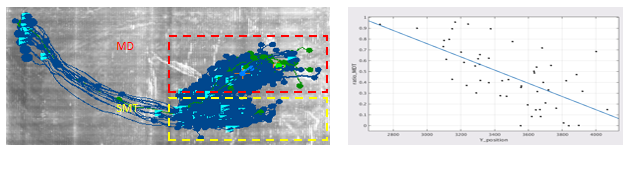
在 CT 神经元中,MD 与 SMT 的投射强度沿背腹方向逐渐变化。
同一神经元类型之间的变异不是随机噪声,而是代表目标大脑区域的偏好
神经元发放模式【树触结构】
不同树突【参与了电信号的整合】位置产生的电信号不同——不同Branch线性相加、相同branch非线性
树突——与神经网络的输入类似;不同信号的整合后——输入到胞体【感觉运动转化等】,越靠近胞体的信号越复杂【不同输入发生了整合】
模型:
1. 单神经元模式:Hodgkin-Huxley model【不同神经元的形态和基因表达不同!因此把每个神经元看成是点对点是片面的】:但解释了动作电位的产生注
神经网络建模
神经元编码方式:Spiking vs Analog【存在域下膜电位】
神经元的形态:轴突越长/胞体越大——树突越大越多;锥体细胞有很多各向异性,并非是用点神经元就可以解释
计算神经生物学 :电信号如何编码(频率、时间、集群)【如何和外界信息对应——如视觉信息,神经元和行为的关联性? 】?信息如何通过突触【是否如电话的中继站,是否只是简单把信号放大】?神经元信号: ^68daaf
突触的信号:对高频信号衰减、对低频信号通透【被动】
Spike【动作电位】:间距有时候密集有时候稀疏,有时候波形也不同【可能和行为产生的外界输入有关】
Na离子和K离子的电流:对膜的穿透存在门控机制(HH model)——v、(m、h、n——状态跃迁过程:开放或不开放,都与电压有关)
不同神经元受刺激时产生不同模式的spike,如Tonic spiking, tonic bursting——依赖于形状和离子通道【因此可基于此对神经元进行分类】
Integrate-and-fire model【HH model无法解释神经元网络,唯一一个模拟大规模的神经网络模型 】:去掉离子电流,只考虑膜电位【当触及阈值,就reset归0】:神经网络中每一个神经的膜电位在随机的触发,每一个神经元的动作电位受到输入影响【影响往上走往下走】,到阈值时说明动作电位发放——分析随时间的up and down。
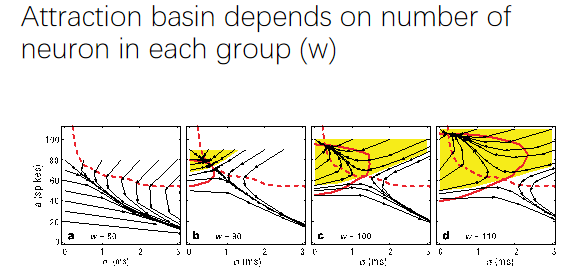
同步化的信息是否真实向下传递?Synfire chain model ——上一层神经元的发放率和同步性会影响下一层神经元的发放率和同步性【a,σ】,最后在虚线上方的会收敛到一个状态【a和σ固定——attractor(吸引子)】,反之在虚线下方的西格玛会变得无穷大,且频率也会变的很小,导致最后一层什么信号都接收不到。【证明了大脑能够通过前馈系统工作 ——即初始频率足够高,可以稳定无损失的把信号传递到最后一级,神经元数量足够多,信号更容易被稳定传下去,即吸引子范围变大】
Wilson-Cowan model:神经系统的状态易发生改变,比如学习的记忆过程,上课后和上课前的状态不是一个


















 2. Cable theory:不同端的树突分成一个电缆【只考虑电荷扩散、不考虑离子通道】
2. Cable theory:不同端的树突分成一个电缆【只考虑电荷扩散、不考虑离子通道】 3. 神经元——简化成卷积【基于神经元输入的权重进行整合】
4. Computational model of a deep layer cortical neuron:树突和胞体是分开的电路
1. 胞体传导到树突的信号变慢
2. 树突复杂度【ρ】,κ【耦合系数:胞体和树突之间电阻的大小:trunk】——耦合系数大,ρ大时会产生burst现象
3. 神经元——简化成卷积【基于神经元输入的权重进行整合】
4. Computational model of a deep layer cortical neuron:树突和胞体是分开的电路
1. 胞体传导到树突的信号变慢
2. 树突复杂度【ρ】,κ【耦合系数:胞体和树突之间电阻的大小:trunk】——耦合系数大,ρ大时会产生burst现象 注:上图所示为Burst现象
注:上图所示为Burst现象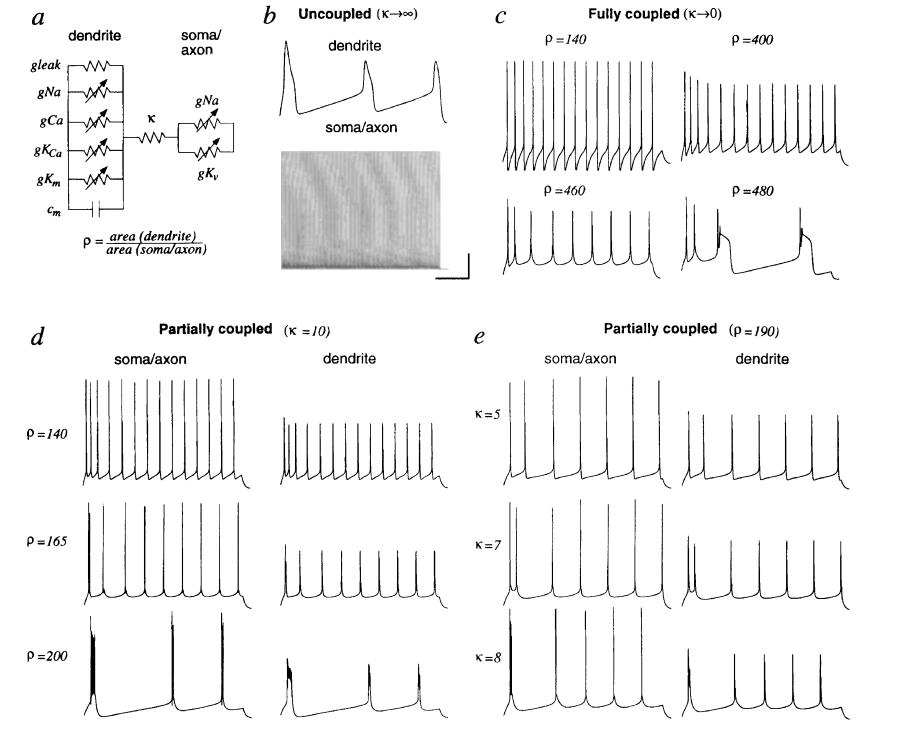 3. 离子通道恒定时,轴突和树突的体积和形态会影响电信号的发放模式
3. 离子通道恒定时,轴突和树突的体积和形态会影响电信号的发放模式