- 新皮质:在哺乳动物之后才出现
- 考题:对未知的内容设计实验流程-有什么方法得到什么结论
- 单细胞转录组测序——不同区的表达谱;不同年龄段的表达谱
- 胚胎电转+病毒转染——示踪(live image)

- 基因功能:胚胎电转做基因的过表达(如把灵长类基因转入小鼠的脑中,产生沟回等)——测量大脑直径变化+大脑的形态学观察+IF染色(不同Marker做形态学的验证)
- 环路研究:病毒示踪,研究基因缺陷时,环路投射的变化情况,结合行为学一起研究
大脑的发育
- 神经上皮细胞:神经管的形成(中间向两端的闭合)
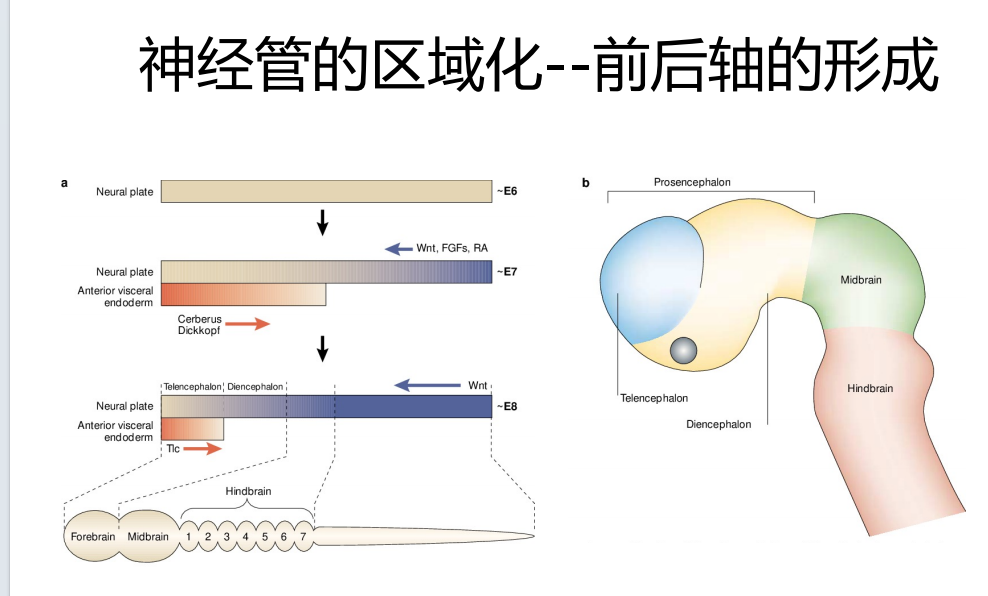
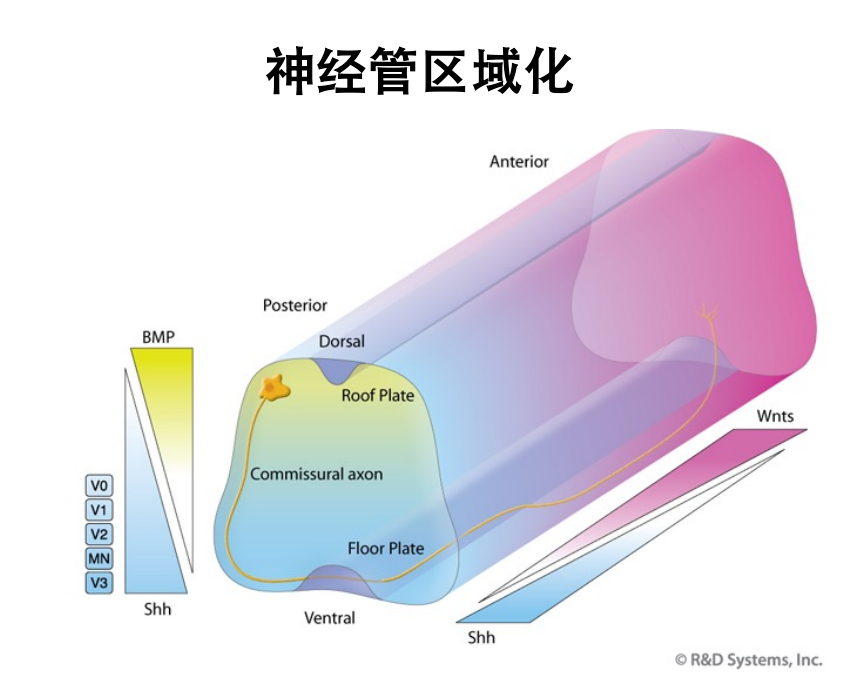
- 神经管的区域化(斑马鱼+鸡胚——再到小鼠的验证):从前到后(wnt、FGFs、RA浓度梯度的形成)+从腹部到背部的(wnt、BMP、shh)复杂过程

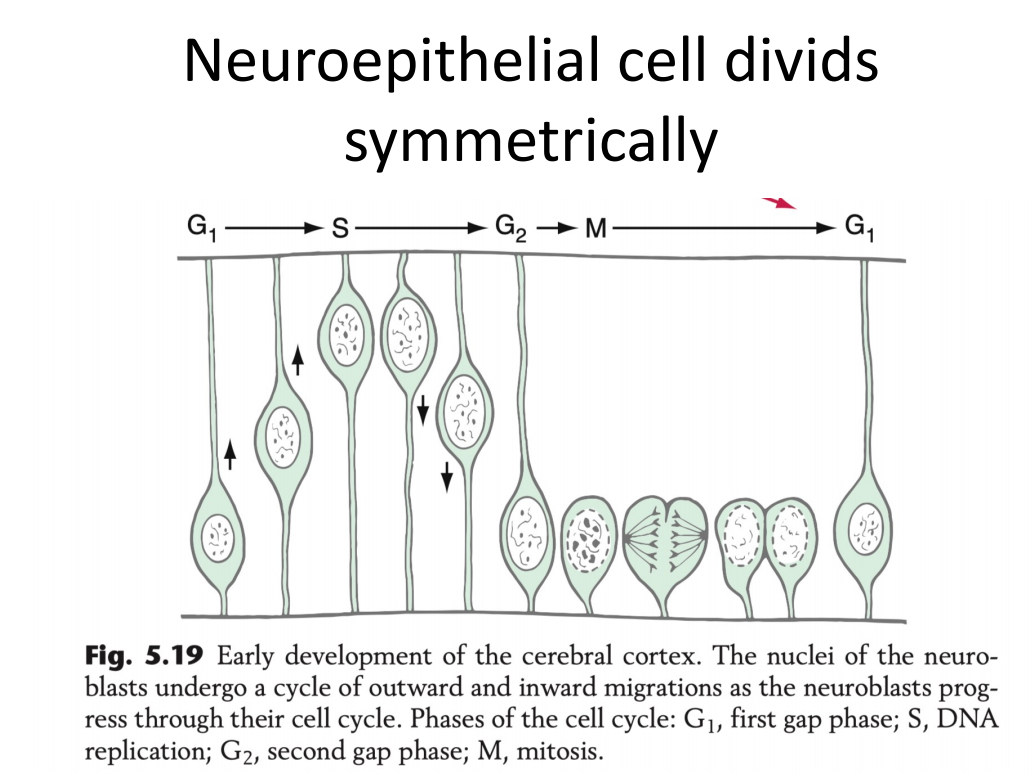
- 大脑的神经发育:神经干细胞在神经上皮出现的核动力迁移的过程(神经上皮细胞的逐渐变厚)
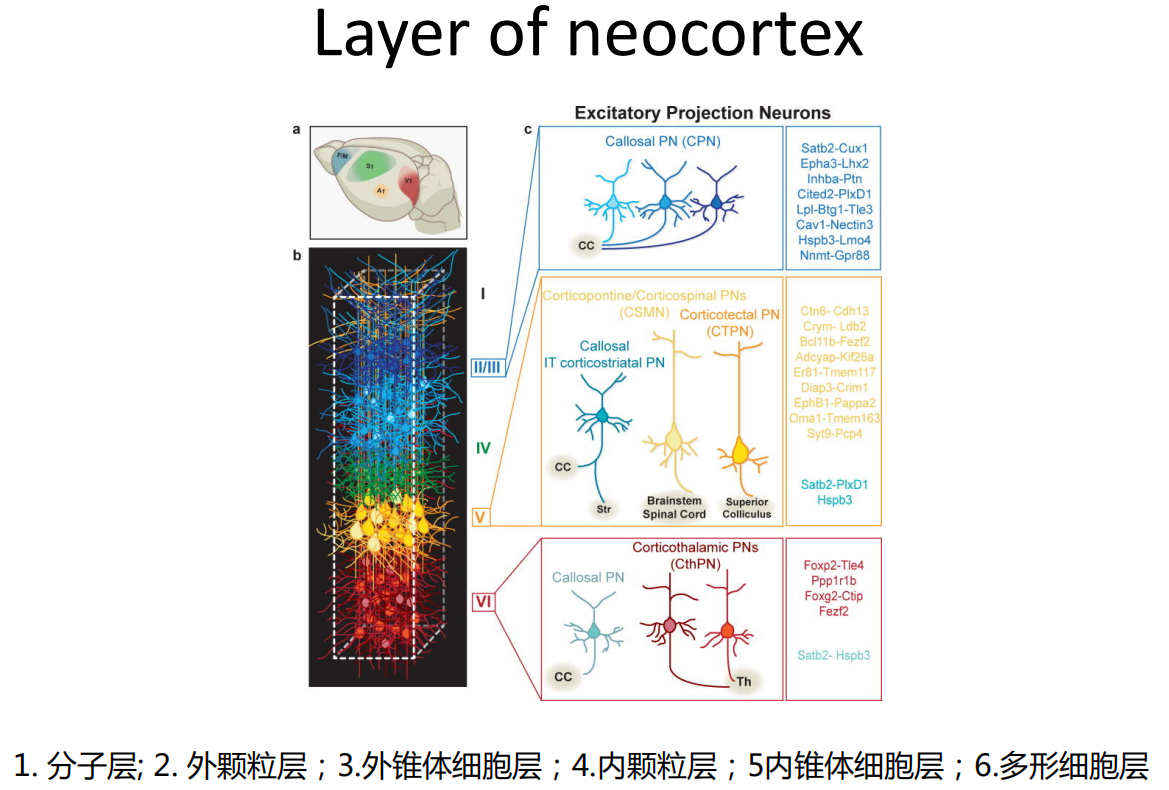 注:发育完成后的大脑皮层
注:发育完成后的大脑皮层 - 神经的发生、迁移和轴突形成
-
Neurogenesis:神经上皮细胞——NE
 注:脑室区的胚胎电转(质粒)和打病毒——进行示踪
注:脑室区的胚胎电转(质粒)和打病毒——进行示踪 -
NE-RGC:这一过程是通过下调上皮相关蛋白的表达(如紧密连接)和上调神经胶质特异性特征来介导的,如糖原颗粒、星形胶质细胞谷氨酸转运体(GLAST)、中间丝波形蛋白,在某些情况下,包括人类,神经胶质纤维酸性蛋白(GFAP)。在这一转变之后,放射状胶质细胞保留了神经上皮细胞的许多原始特征,包括:它们的顶端-基底极性,它们沿着发育中的皮层侧脑室的位置,以及它们的细胞核的阶段性迁移取决于它们与细胞周期的位置(称为“核动力学迁移”)
-
RGC的分裂:
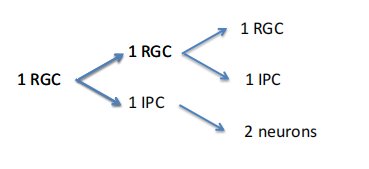
-
oRG cells(可进行多轮分裂,且分裂模式多样无序,maker:HOPX):胞体只能迁移到OSVZ,弹簧式跳跃式分裂(Mitotic Somal Translocation)
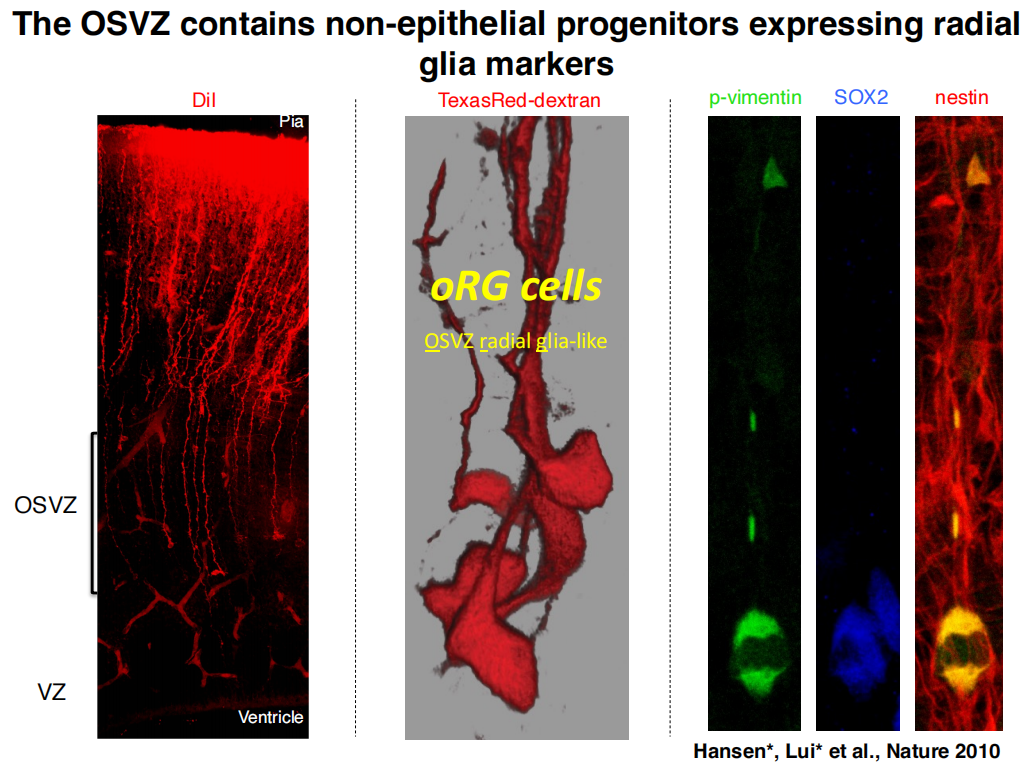
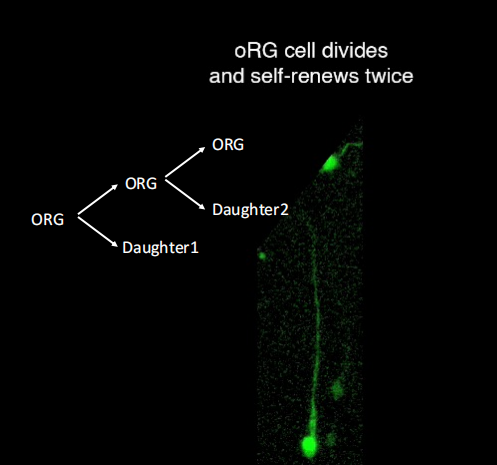
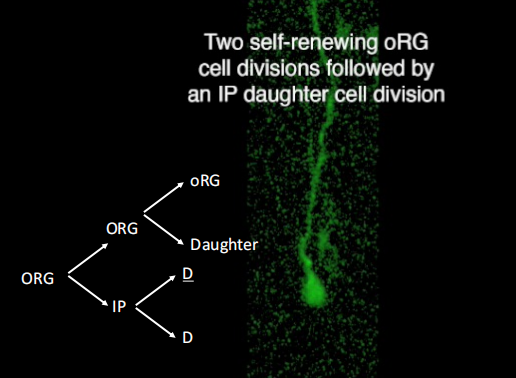 注:oRG细胞的子细胞是神经元祖细胞
注:oRG细胞的子细胞是神经元祖细胞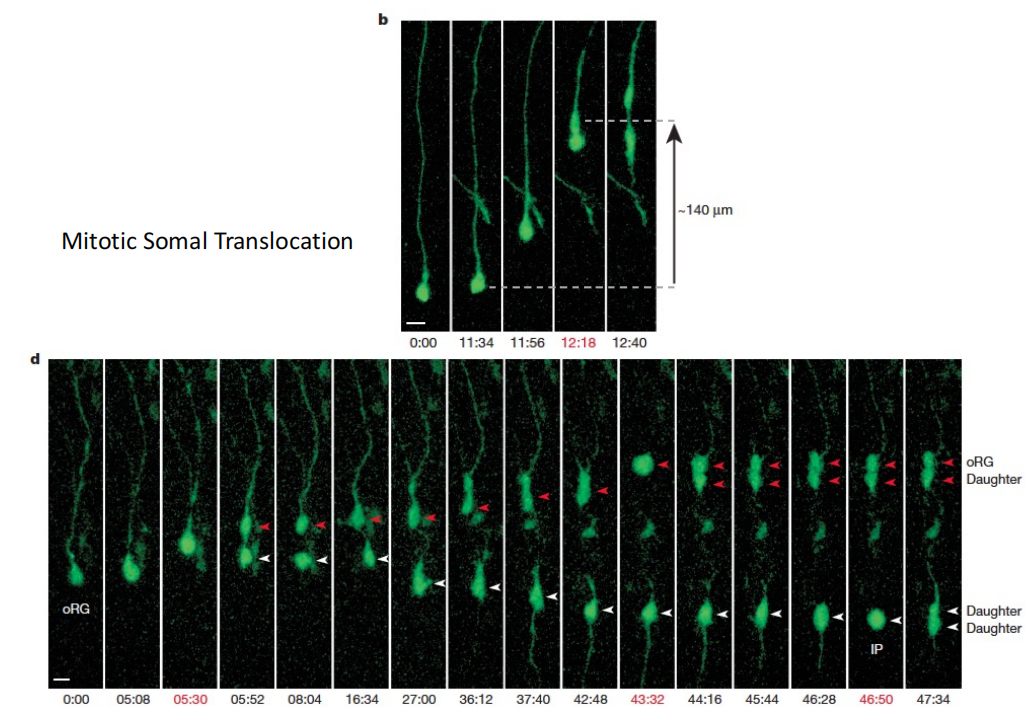
 注:OSVZ并非灵长类动物所特有
注:OSVZ并非灵长类动物所特有 -
VZ区可能形成星胶等可迁移的胶质细胞,OSVZ的RG可能分化出神经细胞等;断层形成白质?
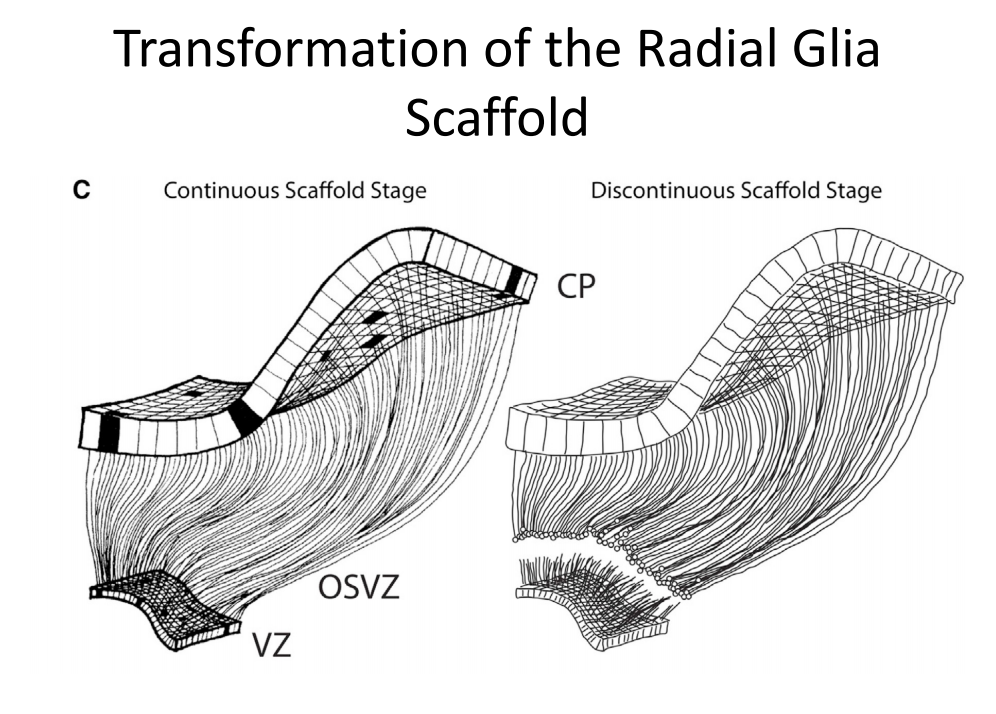
-
总结:
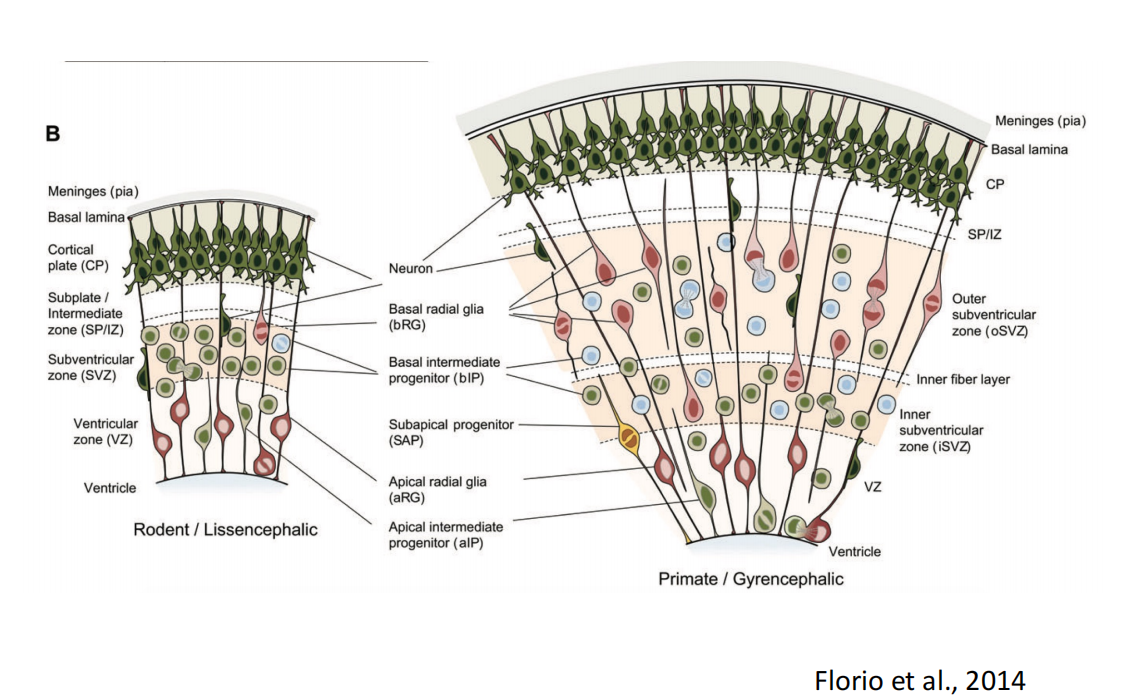 注:(1)IPC细胞90%以上分裂一次(小鼠中),但也可多次(人or 猕猴) (2)OSVZ区以及神经细胞的多少(ORG的存在)不是产生沟回的主要原因
注:(1)IPC细胞90%以上分裂一次(小鼠中),但也可多次(人or 猕猴) (2)OSVZ区以及神经细胞的多少(ORG的存在)不是产生沟回的主要原因
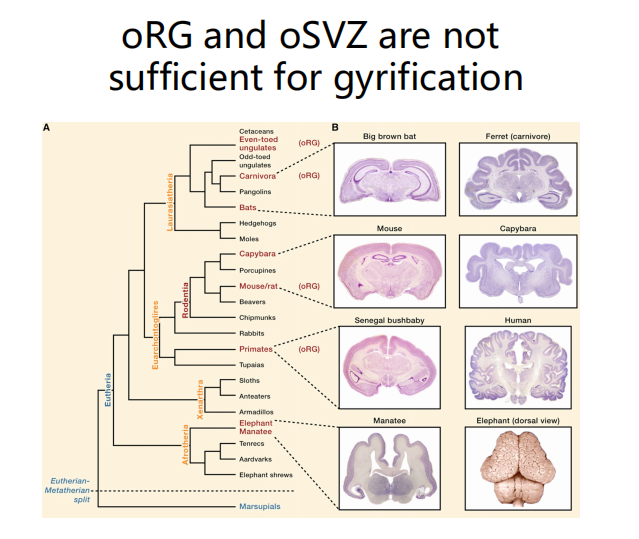 注:沟回或许不是决定智力的因素?
注:沟回或许不是决定智力的因素? -
PCP:神经细胞产生足够多时,干细胞不再产生过多的神经细胞,朝向glia genesis的倾向

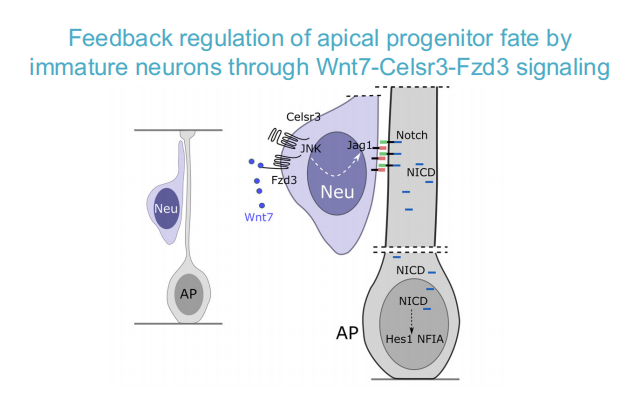
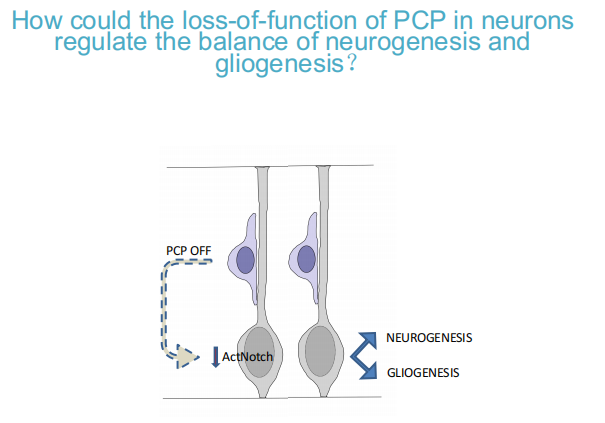
-
兴奋性神经元多产生于背侧端脑,抑制性的interneuron多产生于腹侧:

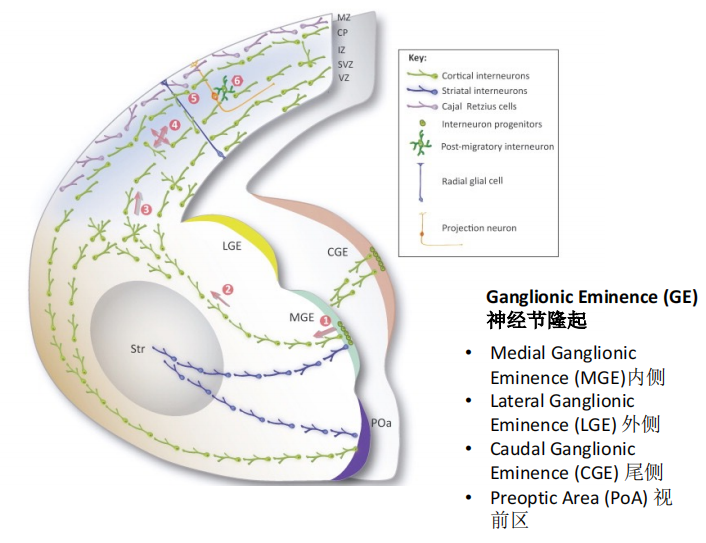 注:GE产生interneuron
注:GE产生interneuron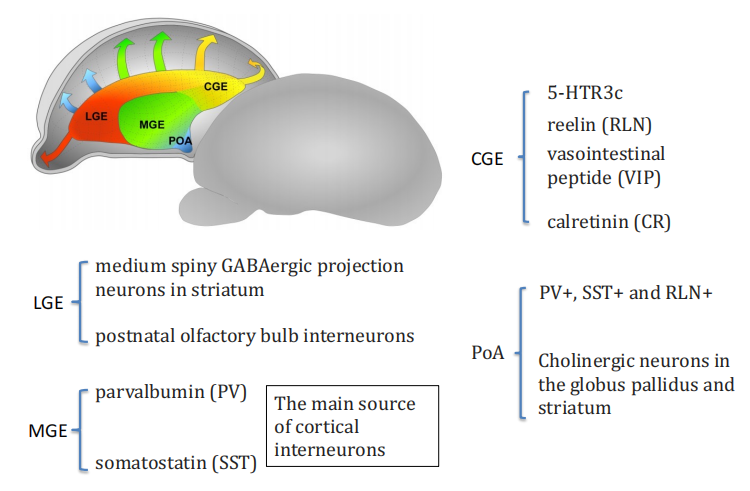
-
Migration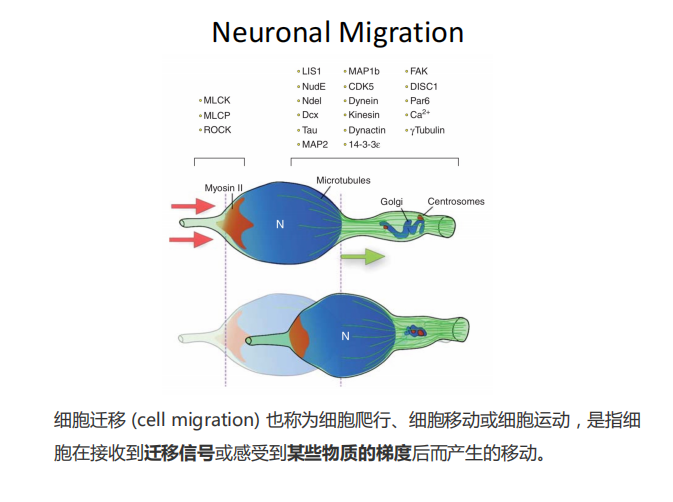
- 迁移方向:radial/横向
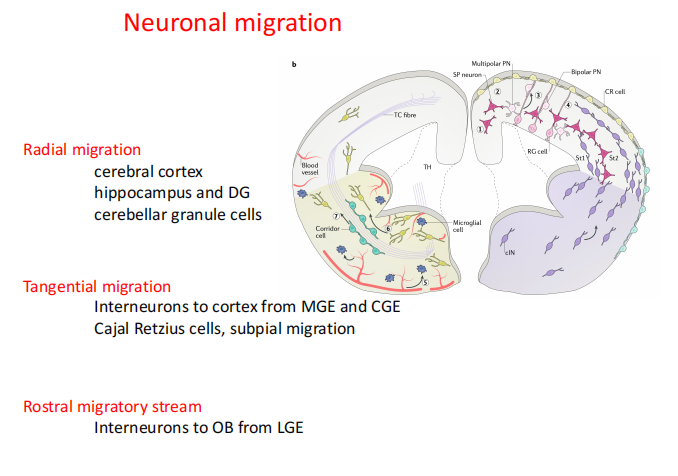
- Radial:
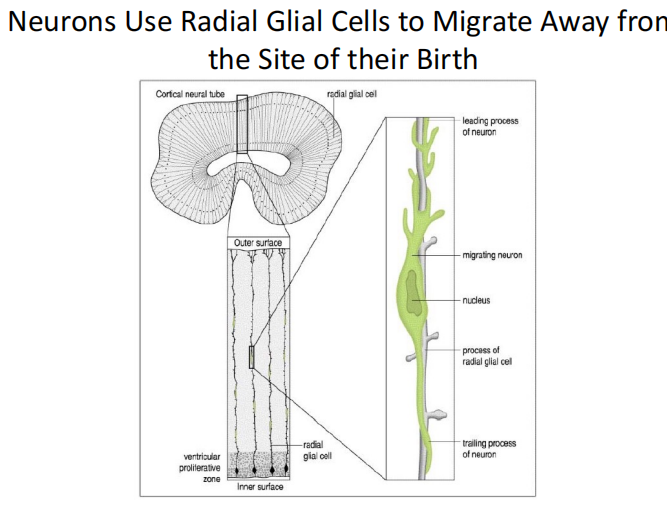 *注:干扰雪貂神经元径向迁移导致无脑畸形
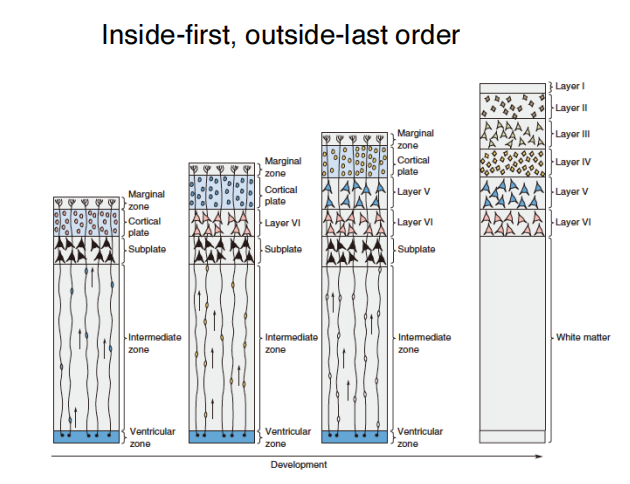
*注:干扰雪貂神经元径向迁移导致无脑畸形 - Inside-out:“后来者居上”
 注:注射BrdU(细胞新生的marker)
注:注射BrdU(细胞新生的marker)
- 疾病模型:
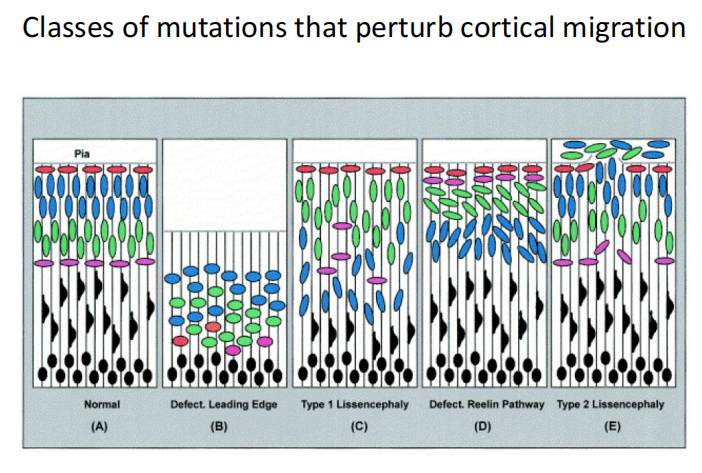 注:人的平滑脑细胞排布是颠倒的过程,即神经细胞排布不同
注:人的平滑脑细胞排布是颠倒的过程,即神经细胞排布不同
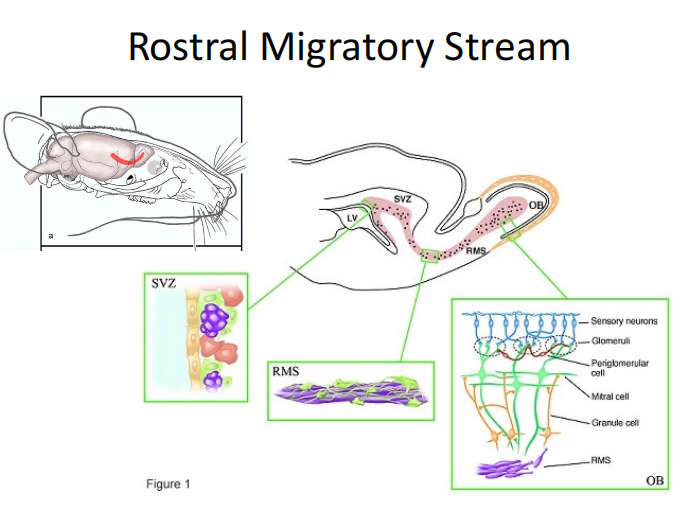
- Rostral migration stream:SVZ区产生的神经细胞沿着RMS迁移到嗅球
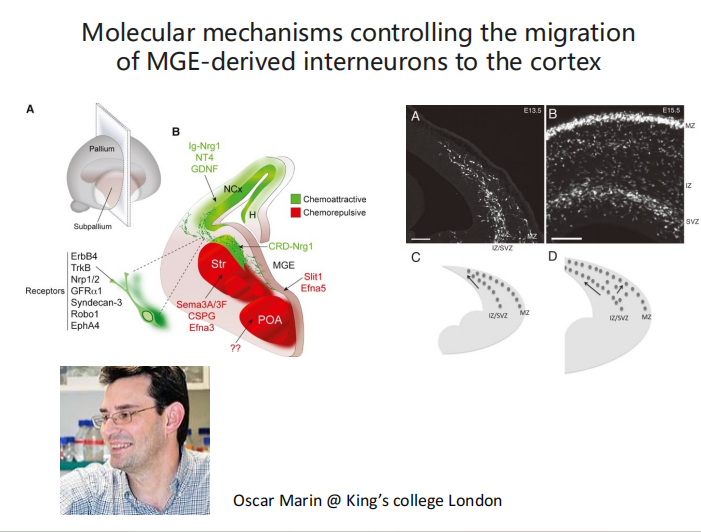
- 切向迁移:与Interneuron迁移有关的信号分子(主要是抑制性的,功能是让神经元的迁移不出错)
- Radial:
Axon growth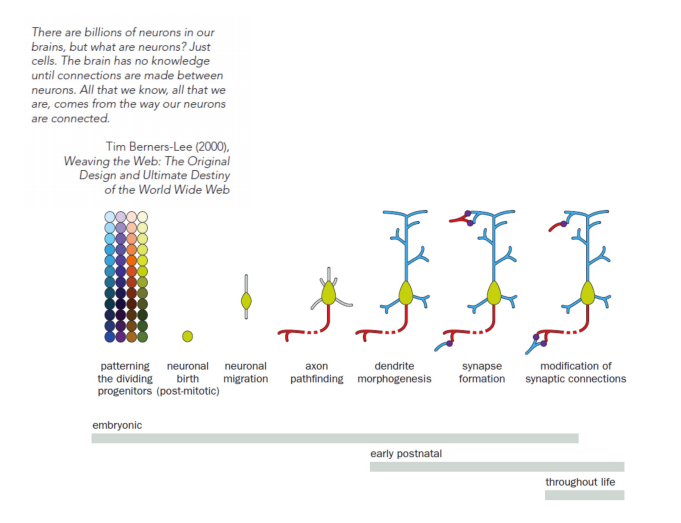
- 神经纤维的分类:
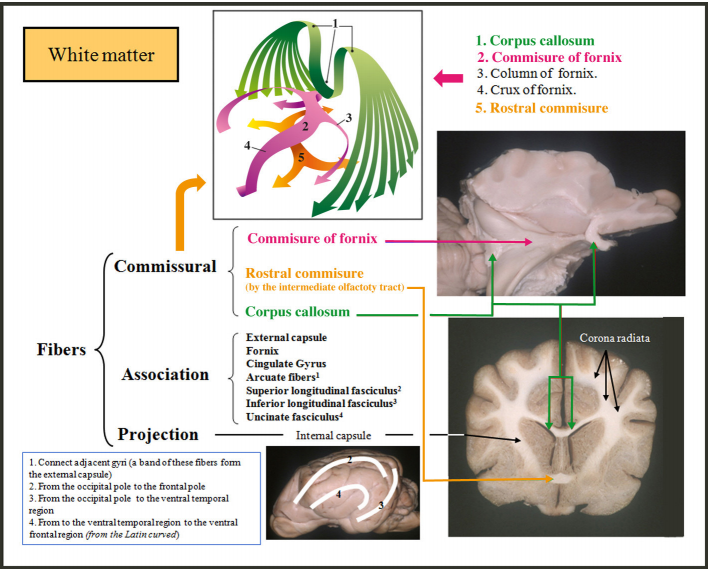
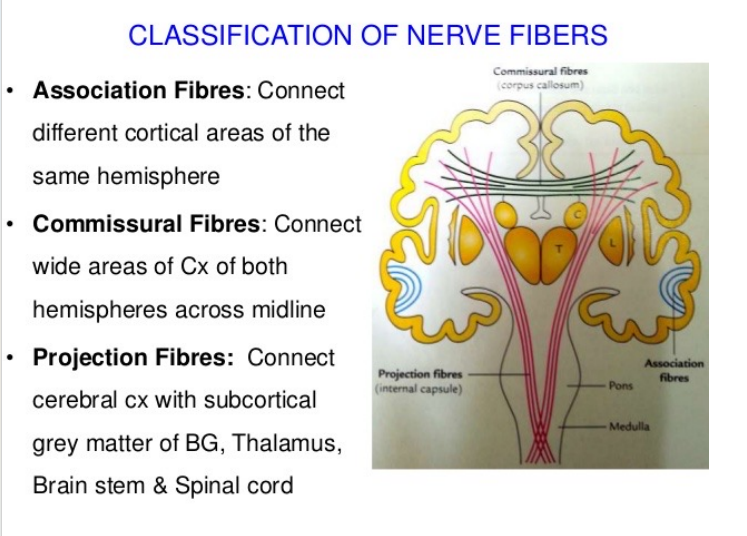 注:胼胝体,最大的白质束,连接两个大脑。
注:胼胝体,最大的白质束,连接两个大脑。
- Axon形成的过程:可受到排斥和吸引信号的影响
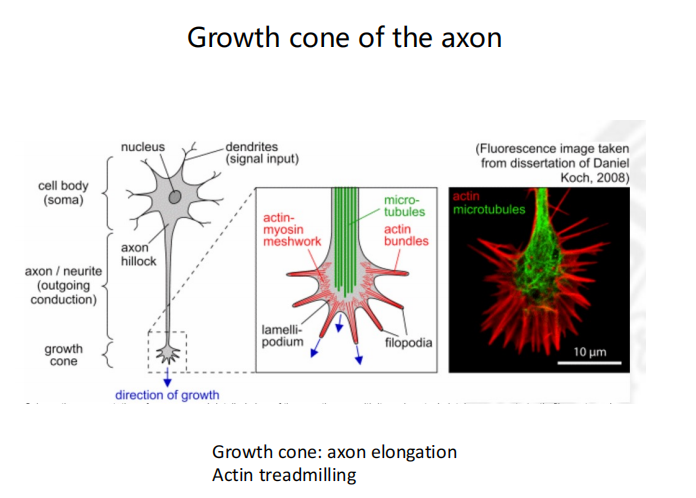
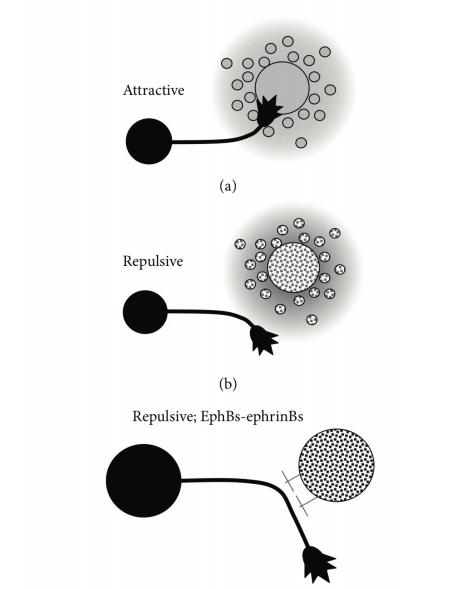

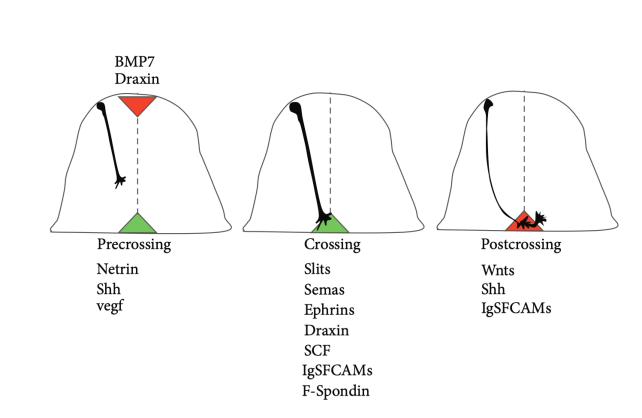
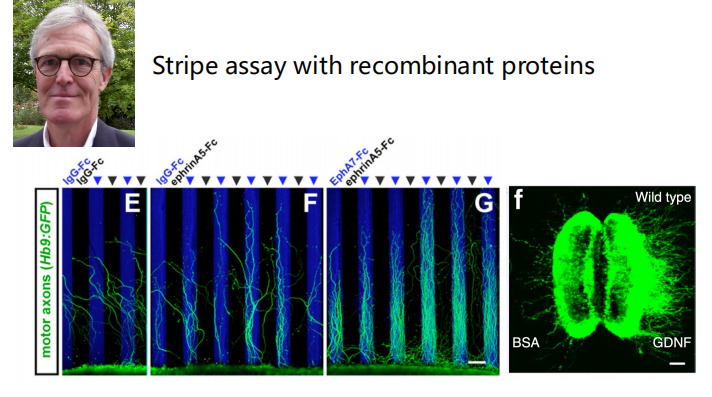
- 研究方法:利用排斥信号和吸引信号结合染色来观察,用重组蛋白进行条纹实验。
- Synapse的形成和修剪

 注:突触的修剪,包括轴突和树突的修剪,是出生后发生的突触消除的过程(~8 mo)•修剪受到环境因素的影响,被广泛认为是学习的代表。
注:突触的修剪,包括轴突和树突的修剪,是出生后发生的突触消除的过程(~8 mo)•修剪受到环境因素的影响,被广泛认为是学习的代表。 - Myelination:
Adult stem cells
- 视网膜祖细胞

- 鸟类:
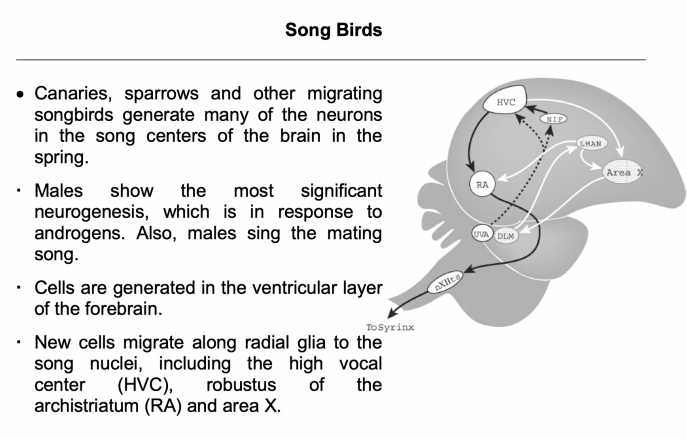
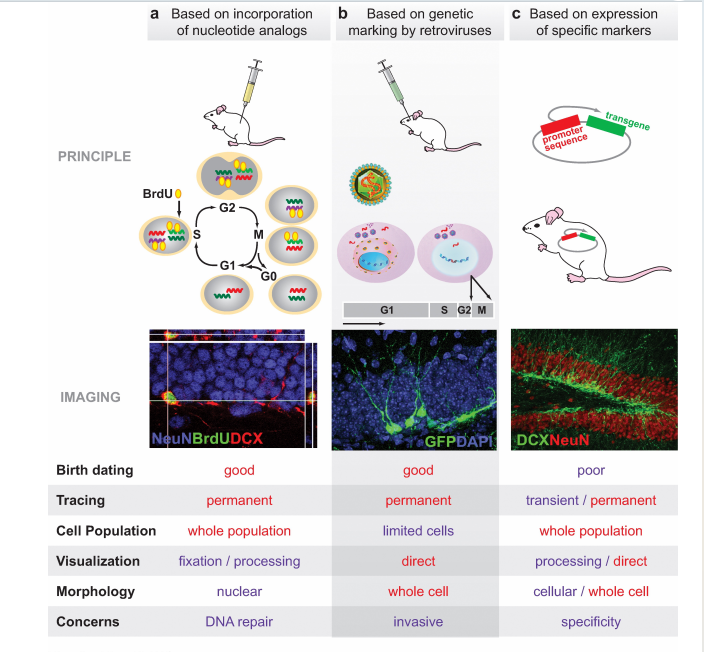
- 研究方法:只有处于cell cycle的细胞才可被标记(BrdU+病毒转染+特定marker)
- 小鼠:

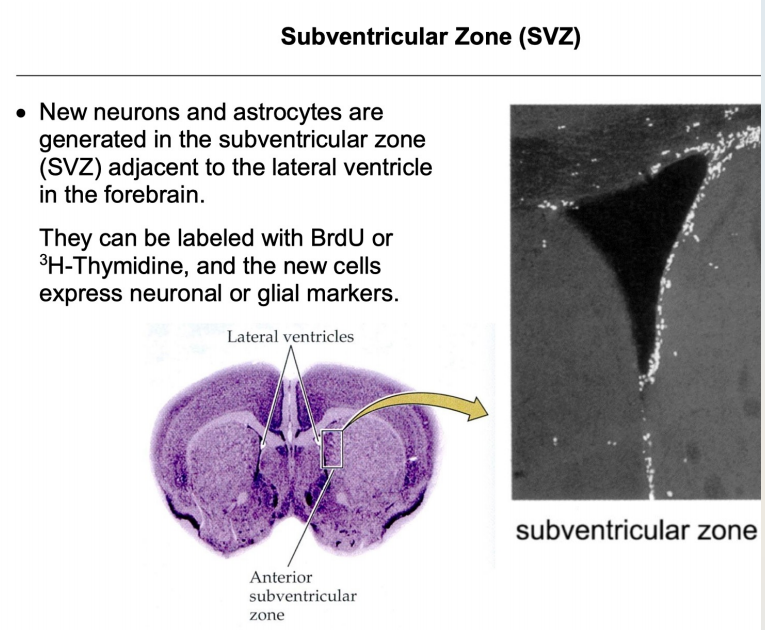
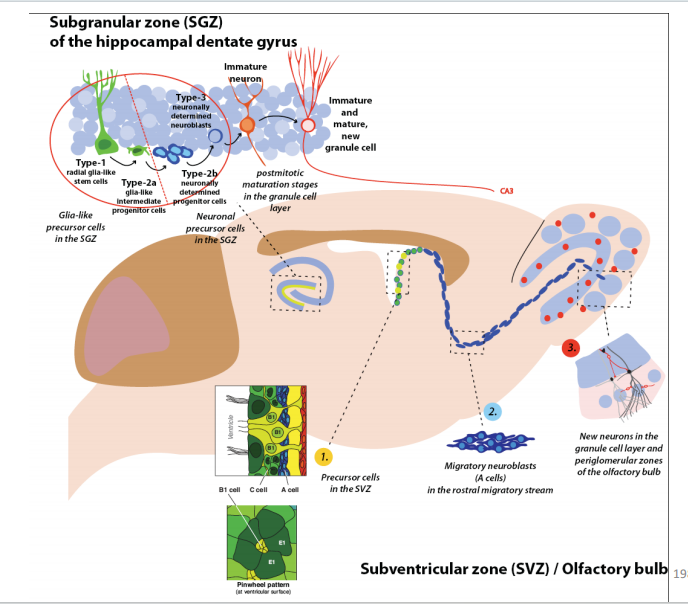
- SVZ:
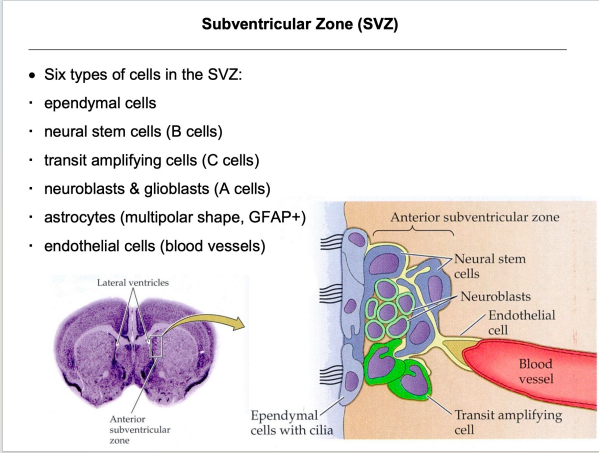 注:室管膜细胞纤毛的摆动机制和PCP有关(不用刻意去记)
注:室管膜细胞纤毛的摆动机制和PCP有关(不用刻意去记)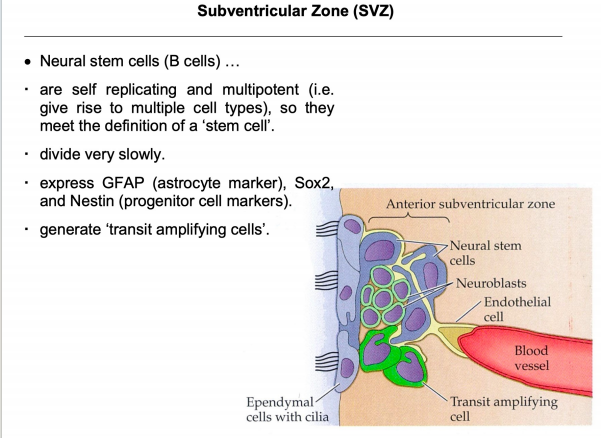 注:成体的Stem cell大部分处于静息状态
注:成体的Stem cell大部分处于静息状态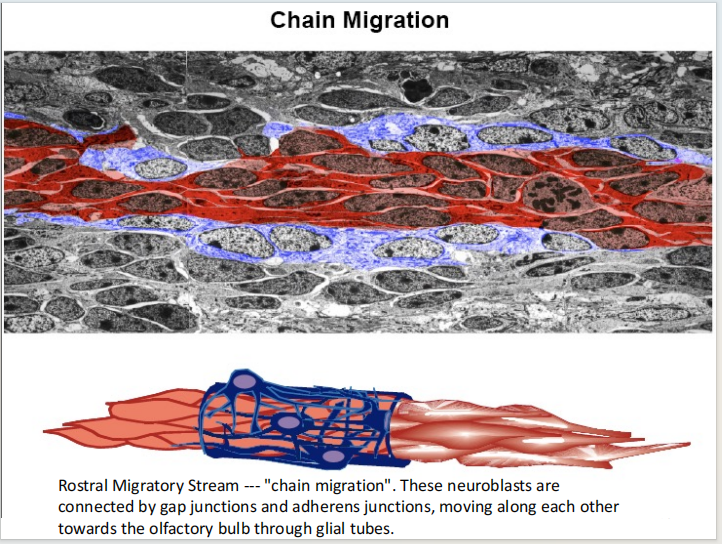 注:RMS式的迁移
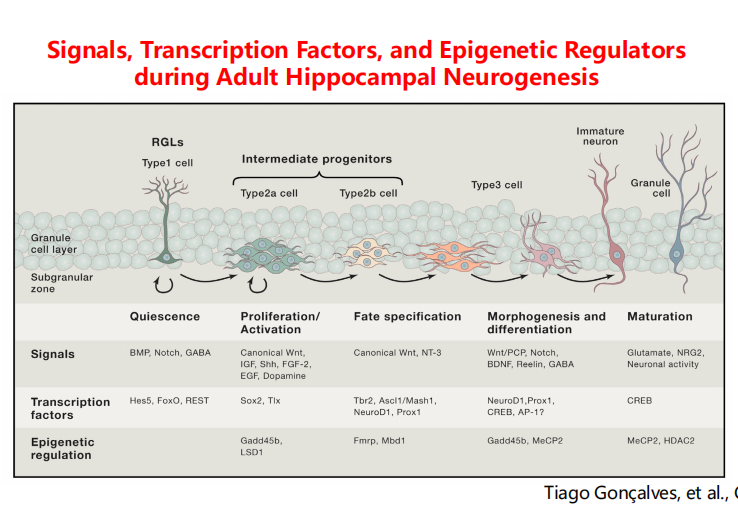
注:RMS式的迁移 - SGZ:学习和记忆的脑区
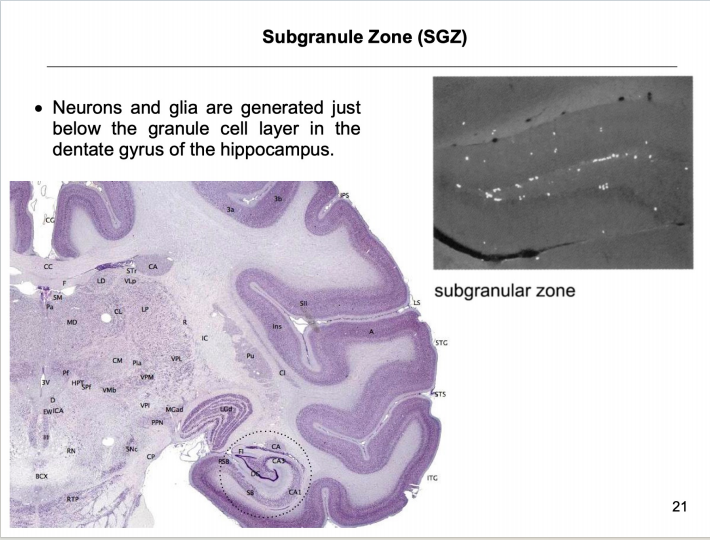 注:颗粒层的下区(发亮点)存在干细胞,神经元和神经胶质细胞刚好在海马齿状回的颗粒细胞层下方产生。
注:颗粒层的下区(发亮点)存在干细胞,神经元和神经胶质细胞刚好在海马齿状回的颗粒细胞层下方产生。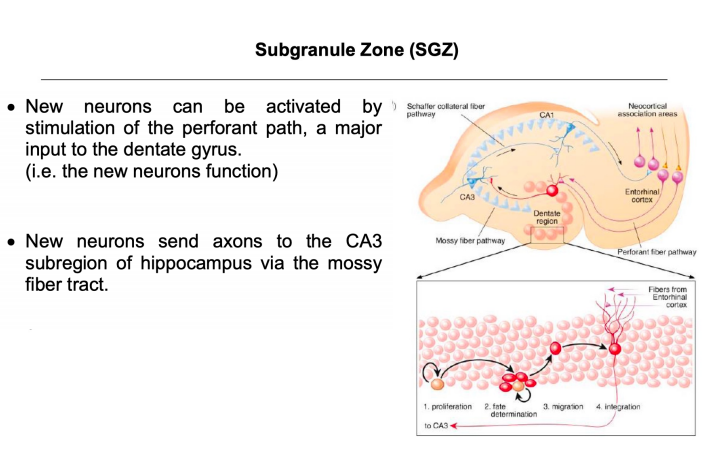
- SVZ:
- 人:有且少,且人和人之间存在差异
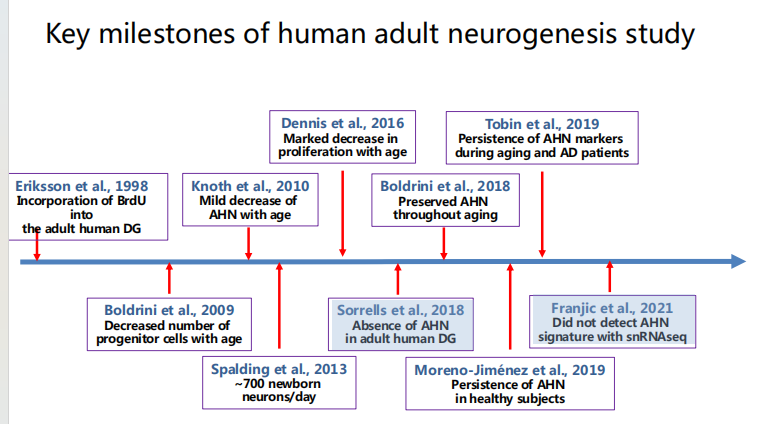 注:为何老年神经发生少?——与个体微环境的炎症程度有关
注:为何老年神经发生少?——与个体微环境的炎症程度有关